
Environmental Setting
I. Estuarine Geomorphology
Excerpted and adapted from the Lake Superior National Estuarine Research Reserve Final Environmental Impact Statement and the Lower St. Louis River Habitat Plan.
The present St. Louis River channel was shaped primarily by the glaciers of the Pleistocene epoch, which began approximately 2 million years ago. As glaciers advanced and retreated, receding for the last time around 10,000 years ago, the melting ice and flowing meltwater left behind complex patterns of sediment, including moraines, drumlins, beach sands, and lake-bottom clays.
Chapter 3: Estuary Characterization
This section covers relevant research on the environmental and socio-ecological settings within the Lake Superior Reserve.
Interactive elements to explore include:
A historic photo viewer contrasting contemporary satellite images of the St. Louis River estuary with the 1861 Hearding Map and aerial photographs from 1939.
The bedrock over which the Upper St. Louis River flows is part of the Canadian Shield, the stable ancient core of the North American continent. The middle river flows over the Middle Precambrian Thomson Formation through Jay Cooke State Park, with whitewater gorges often used for kayaking and rafting. From below Jay Cooke State Park to the Fond du Lac neighborhood, the river crosses the Fond du Lac Formation, which is made up of brown to red sandstone, siltstone, and shale approximately 950 to 1,040 million years old. Below Fond du Lac, coarse grained, dark-gray gabbro forms the high ridgeline on the Minnesota side of the river.

Figure 1: A map of the Pleistocene geology for the Wisconsin Superior region. Credit: Lee Clayton, Geological and Natural History Survey.
This gabbro, along with the fine-grained volcanic basalts that are visible along the Lake Superior shoreline in Duluth, formed as the result of continental rifting about 1,100 million years ago. The St. Louis River below the Fond du Lac Dam is an estuarine system that flows through thick layers of red clay that were deposited approximately 11,000 years ago as the Superior Lobe of the Laurentide Ice Sheet retreated.
After the level of ancestral Lake Superior dropped approximately 1,250 years ago, the river and its tributaries cut deeply incised valleys through the clay. When the lake level rose again, the river valley was flooded, creating a complex estuary with an irregular shoreline and bays at the mouth of each tributary. The ongoing, gradual rebound of the earth’s crust faster to the east and north is causing the water level to continue rising slowly within the estuary. Estimates of the effect of this tilting, called isostatic rebound, on water level at the western end of Lake Superior range from 15cm to 23cm per century.
The large baymouth bars of Wisconsin and Minnesota Points that protect the waters of the Duluth-Superior Harbor are typical of freshwater estuary systems. Historically, there was only one break in the baymouth bar, what is now the Superior Entry, where water from both the St. Louis River and the Nemadji River flowed out into Lake Superior. Completed in 1871, a new channel was dug through the bar on the Minnesota side to facilitate shipping, the Duluth entry. The lakeward side of the bar is composed primarily of sand, and the landward side consists of finer sediments. The baymouth bar shelters the harbor from the wind and wave exposure of western Lake Superior.
Soils within the Reserve are generally clay or riverine deposits and are not well suited for development purposes. Ponding of water, shallow depth to water table, permeability, and shrink-swell properties severely restrict the ability of these soils to support building development, local roads and streets, and underground infrastructure.
II. Climate and Weather
Northwest Wisconsin experiences a cold temperate continental climate that is moderated by the cool waters of Lake Superior. Average annual rainfall is 30 inches and average annual snowfall is 55 inches. Most of the precipitation falls in the summer months from June through September. The coldest month is January with an average high of 20.7°F and an average low of 1.9°F. The warmest month is July with an average high of 76.6°F and an average low of 55.7°F. Wisconsin Climate Change Initiative models predict an increasing frequency of intense storm events, more precipitation falling in the spring and autumn, fewer days of intense cold, and a longer growing season in northwest Wisconsin. Analysis of temperature data from 1950 to 2006 indicate that extreme cold days (temperatures below 0°F) have already decreased by 14 to 20 days and the length of the growing season has increased by four weeks. In the winter, the St. Louis River Estuary is covered by a thick layer of ice that varies from 12 to 35 inches in February depending on temperatures and location. Ice usually develops in mid-November and ice melt usually occurs in late April, although this varies greatly depending on the year.

Figure 2: Annual maximum ice cover across the Great Lakes from 1973 through 2020. Credit: NOAA GLERL and the Wisconsin Initiative on Climate Change Impacts, 2021.
The climate is strongly influenced by Lake Superior, resulting in cooler summers, warmer winters, and greater precipitation compared to more inland locations. Winters are moderately long and somewhat severe, but more than 120 days have temperatures above 50°F (10°C). Average temperatures range from 35° to 50°F (2° to 10°C). A short growing season imposes severe restrictions on agriculture; the frost-free season lasts from 100 to 140 days. Snow usually stays on the ground all winter. The Superior area receives an average of more than 50 inches of snowfall per year.
Exposed coastal areas in this region are subject to significant disturbance from windstorms, waves, ice, currents, and periodic water level fluctuations. These disturbance regimes play a significant role in determining both the landform and vegetation characteristics of the shoreline ecosystems.
III. Hydrology and Oceanography
The hydrologic regime is the primary physical process that determines the morphology and sediment qualities of estuarine aquatic habitats. Water movement through the estuary is complex because it is affected by both the flow of the St. Louis River (and tributaries) and the changing water level of Lake Superior. The water level of Lake Superior fluctuates on a daily, seasonal, and annual basis. Lake level fluctuations lack a predictable pattern and result from the seiche and the variability in precipitation and evaporation, with some influence from the tide.

Figure 3: Historic photo of the falls of the St. Louis River in what is now Jay Cooke State Park, Credit: Whitney Zimmerman.
The bi-directional movement of water acts to both scour areas and deposit sediments. This creates a complex array of protected bays, deeper channels, and shallow flats. The St. Louis River Habitat Plan recognizes ten different aquatic habitat types including natural riverine reach, upper estuarine channels, sheltered bays, clay-influenced river mouths and bays, estuary flats, lower dredged estuarine channels, and industrially influenced bays and slips.
Hydrology also influences nutrient cycling by determining water residence time. Nutrients are carried into the wetlands from three primary sources: 1) the surrounding uplands via the river and its tributaries, 2) water treatment plants, and 3) Lake Superior. Depending on conditions, the St. Louis River Estuary can be either a source or a sink for nutrients. For example, seiche events deliver Lake Superior water high in nitrate into the estuary where summer low-flow conditions increase rates of denitrification, removing nitrate from the system.
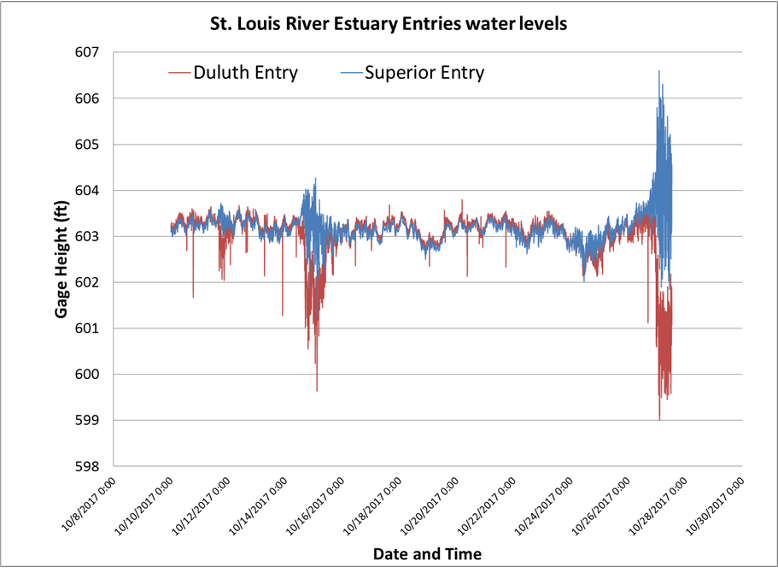
Figure 4: Storm surges, meteotsunamis and other rapid or extreme changes in water levels can be generated by barometric pressure changes, such as this seiche event on October 28, 2017. Credit: Hannah Ramage, Lake Superior Reserve.
The St. Louis River is the largest United States tributary to Lake Superior. As the St. Louis River flows southeast through Minnesota and finally Wisconsin, it encompasses a 3,634 square mile watershed. Near the end of its 179-mile journey through the two states, the river slows and spreads into a 12,000-acre freshwater estuary located in Superior, Wisconsin and Duluth, Minnesota.
The flow of the St. Louis River and fluctuations in water level of Lake Superior determine water level and flow in the estuary. Lake Superior’s water level fluctuates on a daily, seasonal, and annual basis. Long-term lake level fluctuations lack a predictable pattern and result from annual variability in precipitation, ice cover and evaporation.
The hydrology of the St. Louis River is determined by a combination of both surface water and groundwater. Surface water runoff from the St. Louis River watershed is the primary source of freshwater into the estuarine system.
Stormwater management, impervious surfaces, mining, forest management practices, and other land use changes and hydrologic modifications have created water flow that is highly variable compared to pre-development water flow. Today’s early successional forests and deciduous forests do less to slow snowmelt than did the original coniferous forests. As a result, runoff from storms and snowmelt has greater volume, speed, and erosional force, thereby carrying greater sediment loads into these tributary streams and greater erosion of stream banks. Sedimentation is a serious and ongoing threat to the sheltered bay habitats and their accompanying Great Lakes coastal wetland complexes.
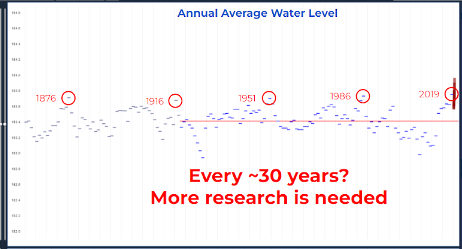
Figure 5: Average annual water level in Lake Superior from 1860 through 2019 shows oscillations on a roughly 30 year pattern. More research is needed to understand the drivers of Lake Superior water levels. Credit: Brandon Krumweide, NOAA.
Groundwater feeds the St. Louis River headwaters, the Seven Beavers/Sand Lake peatlands systems. Despite some early attempts to drain the peatlands and wetlands, much of the peatlands are still present. The wetland areas receive significant contributions from groundwater and contribute to the base flow of the St. Louis River. Natural variability of water flow and sediment transfer into the estuary has also been altered by dams constructed on tributaries as well as the main stem of the St. Louis River. Less than one- quarter of the watershed flow is regulated by the five headwater reservoirs, which reduce natural variability by increasing winter flow, reducing the peak spring run-off flow and severity of flooding, and discharging year-round minimum flows to provide for recreation and fish habitat. The five hydroelectric dams located on the main stem of the St. Louis River have a minimal long-term impact on flows, since they have little storage capability.
IV. Water Quality
The St. Louis River supplies relatively warm, nutrient rich waters to an overall cold and nutrient poor Lake Superior. The seiche and some tidal influence provides the means by which these two bodies of water mix, resulting in the complex water quality dynamics observed in the St. Louis River Estuary. This dynamism contributes to high aquatic biodiversity within the estuarine environment. In addition to seasonal, daily, and even hourly water quality variation caused by climate, precipitation patterns, and seiche events, water quality also varies spatially. In contrast to the thalweg of the St. Louis River, contributions of highly erodible, glacial red silt and clay results in markedly higher turbidity along the Wisconsin estuary shoreline and tributaries including the Nemadji River, Allouez Bay, and Pokegama Bay. These locations also exhibit lower dissolved oxygen concentrations and higher nutrient concentrations than the thalweg (Bellinger et al 2016).
Although current water quality conditions support a healthy ecosystem, the St. Louis River Estuary is one of 43 historically contaminated areas in the Great Lakes (MPCA and WDNR 2013) that was heavily impacted by legacy industrial and municipal contamination. Prior to the passage of the Clean Water Act, raw sewage and industrial wastewater were discharged directly into the St. Louis River Estuary. The resulting high concentrations of phosphorus, ammonia, and total suspended solids led to low dissolved oxygen levels and massive fish die offs. Industrial inputs to the river also included several contaminants such as mercury, polychlorinated biphenyls, polycyclic aromatic hydrocarbons, and dioxins (EPA. n.d.). Deforestation associated with previous logging also contributed to excessive loading of sediments and nutrients. Construction of the Western Lake Superior Sanitary District treatment plant in 1978 increased the treatment standard of municipal effluent, leading to considerable improvements in water quality especially in well mixed portions of the estuary (Alexson et al., 2018; Bellinger et al., 2016; MPCA and WDNR 1992).
Despite these improvements, the St. Louis River Estuary still exhibited elevated total phosphorus concentrations into the early 1990s, necessitating the listing of a Beneficial Use Impairment (BUI) under the Great Lakes Water Quality Agreement for excessive loading of sediment and nutrients (Bellinger et al., 2016; MPCA and WDNR 1992). Water quality data collected at two stations from 1953-2013 under the Minnesota Pollution Control Agency (MPCA) Milestone Monitoring Program was used in conjunction with estuary-wide sampling from 2012-2013 to assess water quality improvements. The findings revealed that there was no hypoxia in the thalweg of the estuary, and that total phosphorus, total suspended solids, and chlorophyll-a concentrations had fallen below the thresholds outlined in the Beneficial Use Impairment (BUI) 6 blueprint, allowing for the removal of the Excessive Loading of Sediments and Nutrients BUI in 2020.

Table 1: Summary of Water Quality Results for Removal of BUI 6 Credit: Bellinger et al., 2016; US EPA Great Lakes Biology Monitoring Program 1996-2015.
Total phosphorus (TP), total suspended solids (TSS), and chlorophyll-a (chl a) were below criteria thresholds outlined for BUI removal in the majority of the estuary’s surface area. Dissolved oxygen was consistently above 5.5 mg/L, indicating no hypoxia.
The MPCA Milestone Monitoring Program has since ended, but water quality measurements continue to be taken at comparable locations by the Lake Superior Reserve under the System-Wide Monitoring Program (SWMP). SWMP data is available at https://cdmo.baruch.sc.edu/.
Table 2 provides summary water quality data collected at four LSNERR SWMP monitoring stations from 2012 through 2021. Phosphate, ammonium, nitrite, nitrate, and chlorophyll-a are determined from grab samples taken on a monthly basis. The remaining parameters are summarized from readings taken every 15 minutes by Xylem YSI EXO2 water quality sondes at each monitoring station. The listed values are based on data collected April - November, and therefore do not encompass winter water quality conditions.
Figure 6 displays the locations of the four LSNERR SWMP monitoring stations in the St. Louis River Estuary.
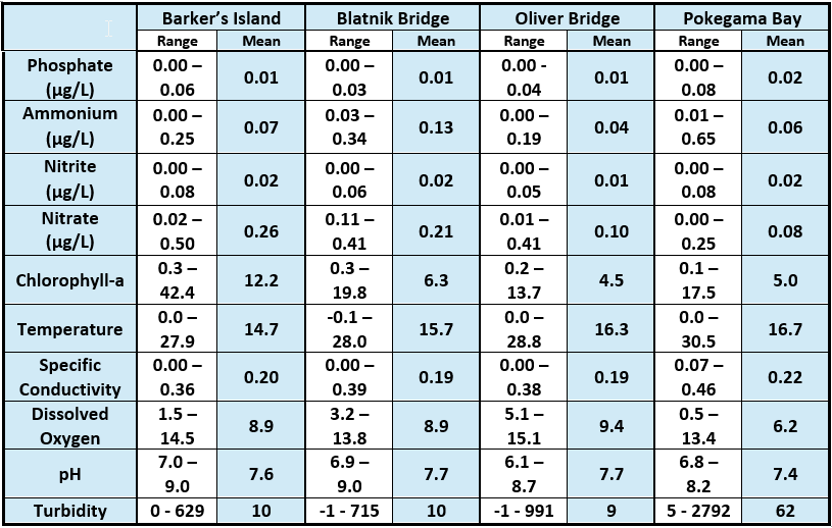
Table 2: Nutrient concentrations and water quality parameters across the St. Louis River Estuary. Credit: Hannah Ramage, Monitoring Coordinator, Lake Superior National Estuarine Research Reserve.

Figure 6: This map displays the locations of the four LSNERR SWMP monitoring stations in the St. Louis River Estuary.
Water temperature is collected from the LSNERR’s water quality monitoring station located at Barker’s Island every fifteen minutes. Annual synthesis of data from May 2013 to October 2021 shows that in 2021, water temperatures were consistently above the nine year monthly averages and were exceptionally high in October (LSNERR 2022).
Recent water quality and phytoplankton research signals rapidly emerging issues, including estuary “hotspots” experiencing cyanobacterial blooms and hypoxia (Reavie 2020). The nitrogen-fixing Dolichospermum (formerly Anabaena) is the dominant cyanophyte in St. Louis River estuary and Lake Superior bloom samples.

Figure 7: Monthly Deviation from Average Temperature at Barker’s Island Monitoring Station Credit: Hannah Ramage and Karina Heim, Lake Superior Reserve.

Figure 8: Harmful Algal Bloom on Barker’s Island, September 2021 Credit: Hannah Ramage, Lake Superior Reserve.
Paleolimnological records indicate that some shallow estuary locales have increasing nutrient loads and cyanobacteria (Alexson et al., 2018). In addition, contemporary observations of harmful algal blooms (HABs) in recreational areas are becoming more frequent (Green 2021). Only recently have unprecedented algal blooms begun occurring within western Lake Superior and its connected waters. In 2021, the Barker’s Island monitoring station was subject to drought conditions and abnormal water quality metrics including high water temperatures, high clarity, and increased productivity.
Reserve staff detected the first confirmed HAB nearby in September of the same year. With limited research so far, Lake Superior blooms appear to be associated with warmer than average water temperatures and may be prone to occur in years with intense storm events that flush nutrients and algal propagules from rivers and estuaries (Reinl et al., 2020; Sterner et al., 2020; Hauser 2018). The nutrient dynamics that facilitate cyanobacterial blooms or HABs within the estuary are still being studied. HABs are triggered by specific environmental conditions and very little is known about the relationships among nutrients, algae quantity, and algal composition in the estuary at this time. Although the St. Louis River is the largest tributary to western Lake Superior, the river’s potential as a source of nutrients and cyanophyte propagules to the Lake are not well understood.
Repeated occurrences of winter hypoxia in multiple estuary locations (Figure 9) were documented in 2013, 2014, 2015, and 2016 (Garono and Schooler 2017). Hypoxic conditions are also known to occur in summer at some locations such as Pokegama Bay, Allouez Bay, and Kimballs Bay. Nighttime dissolved oxygen concentrations at these locations may be low enough to spur phosphorus release from the sediment (Roessler et al., 2018).
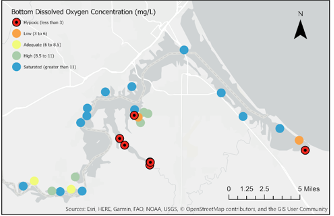
Figure 9: Bottom dissolved oxygen concentrations across the St. Louis River Estuary. Credit: Garono and Schooler, 2017.
Although localized hypoxic events, especially in winter, have been well documented (Garono et al., 2019), no studies have investigated how hypoxic events are triggered or how they impact the ecosystem. In addition to possible storm-related drivers, winter hypoxia in the estuary (Garono and Schooler 2015) may support conditions that foster blooms. Hypoxic benthic environments are well-known to provide ideal conditions for the release of sediment-stored nutrients (Nürnberg 1984), potentially fostering bloom conditions.
Despite remediation and restoration efforts, the St. Louis River and Lake Superior contain elevated levels of mercury and other toxic substances. Consequently, the St. Louis River, including Superior Harbor, is under a fish consumption advisory for mercury in walleye. Fish advisories also exist for high levels of polychlorinated biphenyl (PCB) or pesticides in Lake Superior lake trout, salmon and siscowet. Additionally, the St. Louis River is listed as impaired under state 303(d) lists in both Minnesota and Wisconsin for contaminated sediment, chronic toxicity, and contaminated fish tissue. These contaminants necessitate a BUI for restrictions on dredging activities.
Contaminated sediments contribute mercury to the water column. The most problematic areas of persistent contaminated sediments, such as the industrial slip in the harbor, are located outside of the present NERR boundaries. Isotope analysis has revealed that legacy mercury contamination in the estuary actively bioaccumulates in game fish (Janssen et al., 2021). Per- and polyfluoroalkyl substances (PFAs) are a emerging contaminant of concern; they have been found in smelt in high enough levels to cause a consumption advisory (WDNR 2021).
Socio-Ecological Setting
I. Demographics and Population Trends of Reserve Connected Communities
Prior to European settlement, the region was home to the Fond du Lac Band of Lake Superior Chippewa (Ojibwe) and remains so today, with tribal reservation lands adjacent to the City of Cloquet approximately 20 miles west of Duluth, Minnesota. European settlement and development of the area began in earnest in the mid-19th century. The population of the Twin Ports peaked in the first half of the 20th century, and has since declined. In 2020, the population of Douglas County was 44,295, and its urban center, Superior, Wisconsin was 26,751. Comparably larger are the populations of St. Louis County (200,231) and its urban center of Duluth, Minnesota (86,697). Today, the population of the region is predominantly white. Ninety-three percent of the combined populations of St. Louis and Douglas counties identify as white, three percent identify as American Indian or both white and American Indian, two percent identify as black or African American, and just over one percent identify as Asian.
The Twin Ports communities of Duluth and Superior are an urban metropolitan center and the population hub for the region with a combined 48,252 households. The median household income in Superior ($48,830) and Duluth ($54,084) falls below each community’s respective county and state rate. The housing stock of both communities is majority owner-occupied (56% of Superior homes, and 60% of Duluth homes).
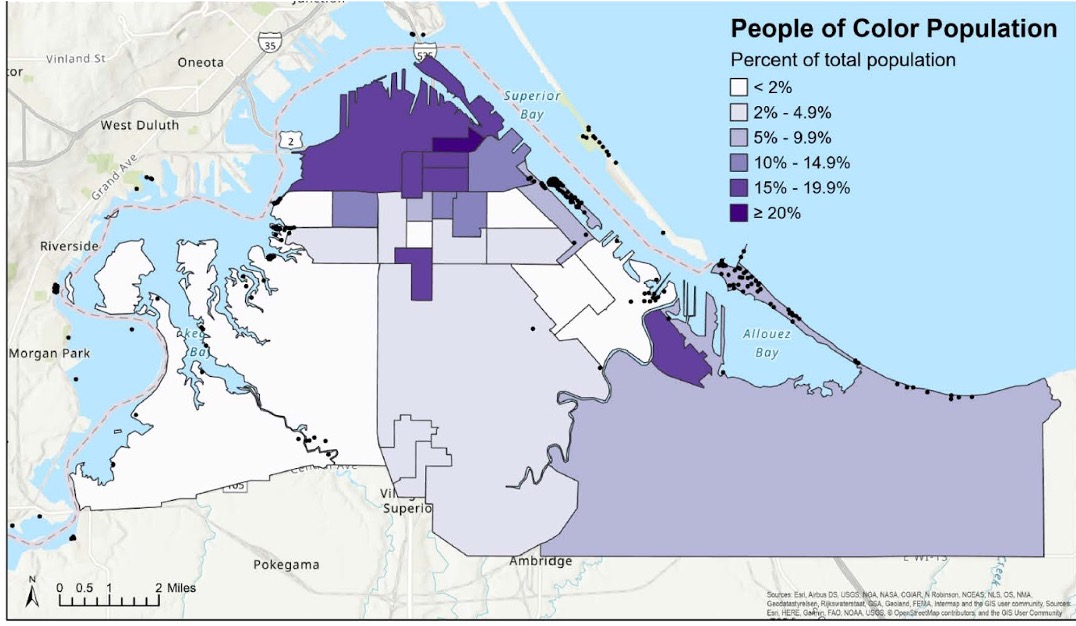
Figure 10: Although 90 percent of the population of the City of Superior identifies as white, this map shows some of the geographic distribution of the non-white population within the city by Census block group (2019 American Community Survey data). Credit: Rebecca Joseph, NOAA Hollings Scholar, 2021.
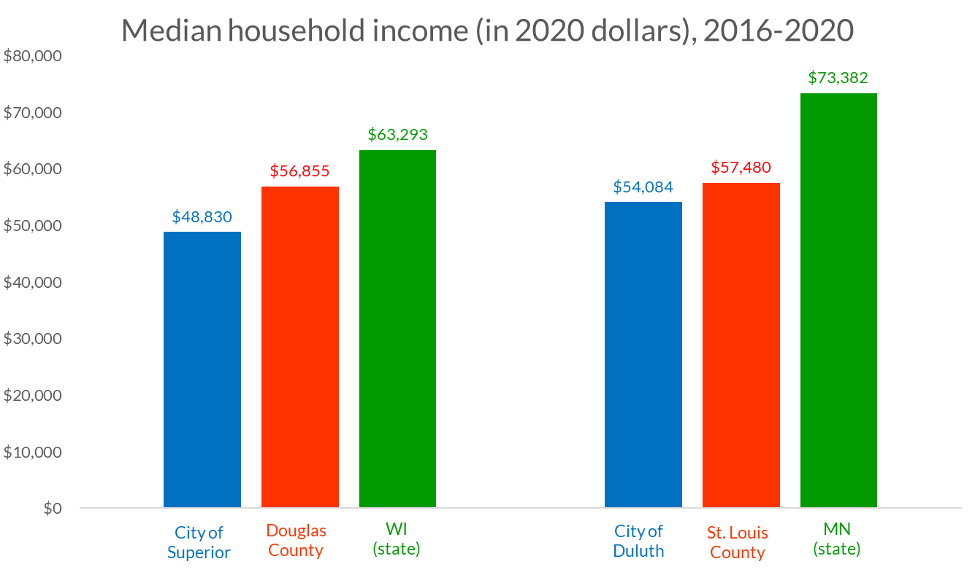
Figure 11 Credit: Lake Superior Reserve Management Plan, 2020.
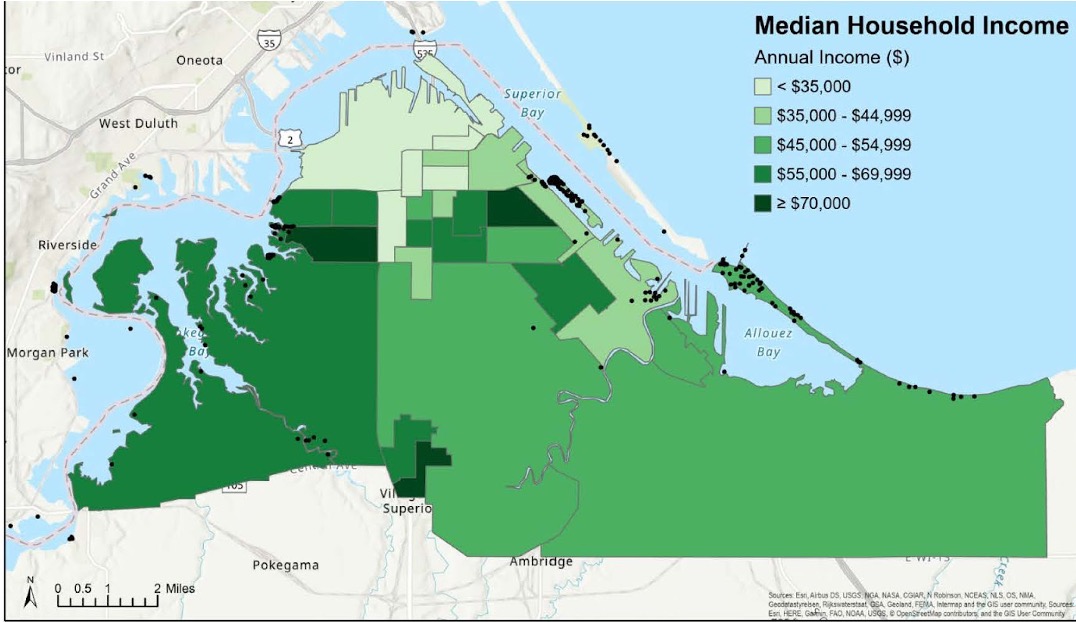
Figure 12: Geographic distribution of median household income within the city of Superior by Census block group (2019 American Community Survey data). Credit: Rebecca Joseph, NOAA Hollings Scholar, 2021.
Educational attainment levels for adults in the Twin Ports region are consistent with state levels. In the region, 94 percent (Douglas County) and 95 percent (St. Louis County) of the population over age 25 have received a high school diploma or high level of education. There are a concentration of universities and colleges in the Twin Ports, and a significant proportion (14 percent) of Duluth and Superior residents are enrolled in an undergraduate college program. In the City of Superior, 54 percent of the population over 3 years of age is enrolled in K–12, and in Duluth, 40.5 percent of the population over age 3 is enrolled in K–12.
II. Ecosystem Services
Much of the St. Louis River Estuary and its surrounding lands stand in stark contrast to the dense urban environment of the cities on either side. Consequently, the estuary is a popular location for recreational activities such as hiking, foraging, hunting, fishing, and boating.
Written by Noah Pinnsonault
Noah is the Lake Superior Reserve Research and Monitoring Technician.
The estuary is also part of the 1842 and 1854 Ceded Territory of the Ojibwe people, also known as Anishinaabe or Chippewa. Ojibwe retain rights to hunt, fish, gather, and practice traditional lifeways within Ceded Territory boundaries. The St. Louis River is also vital to the local economy, of which maritime shipping and tourism are major components. The St. Louis River watershed provides an estimated $5 billion to $14 billion in ecosystem services annually (Fletcher and Christ in 2015).
Trails provide access to the many distinct habitats found around the estuary. The Superior Municipal Forest State Natural Area provides motorized vehicle, hiking, skiing and mountain biking trails around Pokegama Bay, an area of forested wetlands, clay flats, and uplands. Swimming is a popular activity on the 10 mile beach which stretches across Minnesota Point and Wisconsin Point. The Minnesota DNR and the Minnesota Audubon Society recognize the St. Louis River estuary as an Important Birding Area (IBA).
Wild rice (manoomin, Zizania palustris) was historically abundant in areas such as Allouez Bay, Spirit Lake, and in the sheltered bays created by the islands of the upper estuary. Its harvest was a major livelihood activity for the Ojibwe people. Industrial impacts and hydrology changes as well as an increased population of herbivorous Canada geese have reduced the prevalence of wild rice within the estuary. Though the plant retains its central importance within Ojibwe culture across the region, little is harvested from the St. Louis River. An ongoing wild rice restoration project provides new opportunities for cultural revitalization as well as recreation for non-Indigenous residents.
Black ash (baapaagimaak, Fraxinus nigra) is used extensively in traditional Ojibwe basket and tool making. The tree is abundant on islands within the estuary as well as the surrounding lands. However, the future of this craft is threatened by the invasive Emerald Ash Borer (Agrilus planipennis), which was confirmed to be present in Superior, Wisconsin in 2013 and has spread widely since. Other plant species, such as ramps or wild leeks, blueberries, strawberries and medicinal herbs are gathered nearby. Tapping trees for sap, primarily maple for the production of syrup and maple sugar, is also a common practice on the forested lands adjoining the estuary.
Gun hunting and archery hunting are popular in areas such as the Pokegama Carnegie Wetlands, Superior Municipal Forest, and the St. Louis River Stream Bank Protection Area. Prey animals include white-tailed deer, black bear, ruffed grouse, duck, goose, turkey, and more. The presence of fur-bearing animals such as beaver, otter, muskrat, fox, and coyote makes these locations attractive to trappers as well.
Advisories limit the number of fish eaten from the St. Louis River. Concerns include high levels of mercury in walleye. Lake Superior lake trout, salmon, and siscowet are also under fish advisories for levels of polychlorinated biphenyl (PCB) or pesticides. High concentrations of PFAs are problematic in smaller fish such as smelt (WDNR, 2021). Despite this, water quality improvements and habitat restoration have precipitated an increase in the popularity of fishing on the estuary. It is inhabited by over 45 native species of warm water fish, including walleye, channel catfish, northern pike, muskellunge, smallmouth bass, black crappie and lake sturgeon.
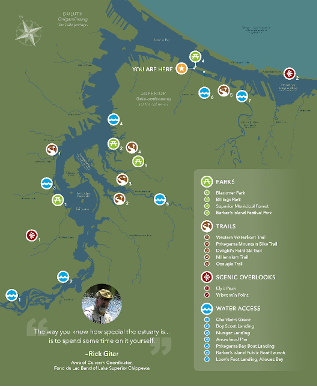
Figure 13: Access map of the St. Louis River estuary developed for the Lake Superior Estuarium. Credit: Sarah Congdon Lake Superior Reserve.

Figure 14: Smelt are typically caught using seine nets along Lake Superior beaches in early spring. Credit: Deanna Erickson, Lake Superior Reserve.
Netting of the non-native smelt (smelting) is a common occurrence in the spring. While some fish from shore on public docks, fishing from boat is made possible by the numerous public watercraft accesses around the estuary. The public accesses are also used for launching kayaks, canoes, and other recreational watercraft onto the St. Louis River Estuary National Water Trail that winds through the upper estuary.
Maritime commerce is the dominant industry on the Lower St. Louis River Estuary. The Duluth-Superior Harbor spans an area of 19 square miles and 49 miles of shoreline. 35 million short tons of cargo pass through the port on a yearly basis. Primary cargoes are iron ore, coal, limestone, and bulk grain. The port necessitates the maintenance of 17 miles of dredged channels.
III. Culturally Significant Sites and Human Dimensions Within the Reserve
Culturally Significant Places
The estuary as a whole is a culturally significant place to both Indigenous and settler people of the region. Historically, the Dakota and Ojibwe cohabited the area for a time and also came into conflict over territory, including a battle on Rice’s Point, approximately 5 miles upstream from the mouth of the St. Louis River. The region is a continuous home to the Fond du Lac Band of Lake Superior Ojibwe (Chippewa), with reservation lands located in the St. Louis River watershed, and both ceded and unceded lands in Superior and Duluth.
Excerpted and adapted from the Lake Superior Management Plan.
In Anishinaabe migration stories, Manidoo-minis (Spirit Island), located in Chi-gami ziibi (the St. Louis River) became a stopping place along the transcontinental migration of the Ojibwe people, where the people were directed to “the place where food grows out of the water.” Spirit Island is extremely important to the many bands that make up the Anishinaabe Ojibwe Tribal Nations. The island was regained by the Fond du Lac Band in 2011 through purchase. Burial mounds exist throughout the region. A cemetery, primarily of Ojibwe people, existed on Wisconsin Point in the village of Chief Joseph Osaugie and his descendants. Many bodies were disinterred and moved by garbage scow to St. Francis Cemetery in Duluth in 1914. In 2022, the City of Superior formally returned the burial sites to the Fond du Lac Band of Ojibwe.
Anishinaabe encampments, village sites, cemeteries and ceremonial areas were and are located around Spirit Island, including Minnesota Point, Wisconsin Point and Nekuk (Nigig, otter) Island in the upper estuary. The productive and expansive estuary contained abundant fish, game, wild rice and waterfowl, and served as an important center of fur trade corridors up to Knife Portage (now in Cloquet, MN), as well as at both ends of the Nemadji River. Nagaajiwanaang, the Fond du Lac village at the base of a series of falls and now the Fond du Lac dam, was the gateway to Sandy Lake and the Mississippi River by way of the St. Louis River, with a nine-mile portage as the only means of access. Wild rice continues to be of seminal significance to Ojibwe and other Tribal Nations of the region and its restoration in the estuary is an ongoing priority for the Indigenous and land management community. Fire scars across the region indicate that areas forested with pine, including Wisconsin and Minnesota Points, were likely stewarded through cultural burning to promote the growth of blueberries and other plant medicines.
European exploration and eventual settlement of the St. Louis River began in the mid 17th century by French fur traders and missionaries. By 1787, the North West Fur Company, an association of Montreal merchants, established trading posts in the estuary with a permanent post (Fort St. Louis) built at Connor’s Point in what is now Superior, Wisconsin by 1793. In 1816, a combination of events including competition from other fur trading companies and the War of 1812 led to the closing of the post and its re-establishment in Fond du Lac, MN. While the fur trade continued until the 1870s, the post itself closed in 1847 due in large part to the declining human and fur-bearing animal populations brought on by Eurasian diseases and overexploitation.
Human Dimensions
The Lake Superior Reserve is situated near or within the Twin Ports cities of Duluth, Minnesota and Superior, Wisconsin, which have become a regional retail and service center for northern Minnesota and northern Wisconsin. Tourism and recreation drive the Great Lakes economy in the Twin Ports, with some 6.7 million tourists visiting each year, drawn in large part by the beauty and natural amenities of the St. Louis River and Lake Superior. Multiple universities and colleges in the region, a large industrial port, two large medical facilities, energy and trade, as well as education and health services, contribute significantly to the overall coastal regional economy.
The coastal economy of St. Louis and Douglas counties comprises all activities and industries reported by the Bureau of Labor Statistics. In 2015, the Natural Resources and Mining sector and the Trade, Transportation, and Utilities sector were the largest in terms of their contribution to the combined gross product of the two counties, each contributing 17 percent of the total. In contrast, the Education and Health Services sector is the largest across the two counties in terms of employment numbers, making up 33 percent of coastal employment, followed by the Leisure and Hospitality sector with 12 percent.
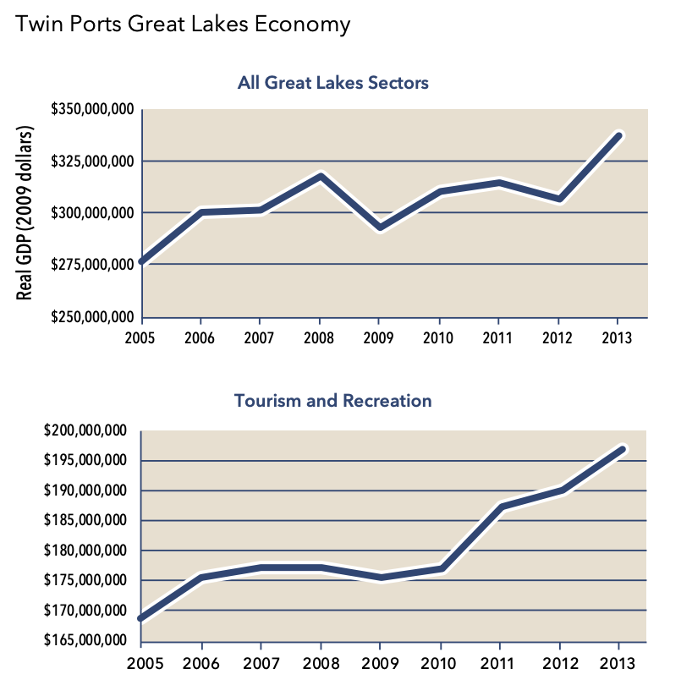
Figure 15: Graphs depicting 2009 Real GDP across 1) all Great Lakes economic sectors, and 2) specific to the Tourism and Recreation sector.
The Great Lakes economy includes only those sectors and industries which are closely related to marine activities, as defined by the NOAA Economics: National Ocean Watch. The Great Lakes economy in Douglas and St. Louis counties, defined in terms of gross product, was impacted by the 2008 financial crisis but has been recovering since with significant growth starting in 2012 and extending through the most recent data year in 2019. Jobs in Great Lakes economic sectors have also been rising since the mid-2010s. From a recession-era high of 10 percent in 2009, the unemployment rate in the Duluth-Superior metropolitan area had been on a steady decline until the COVID-19 pandemic began in 2020.
Social Vulnerability
The Lake Superior Reserve has a relatively high social sensitivity index value, according to the 2013 Climate Sensitivity of the NERRS report. This report indicates a variety of reasons for higher social sensitivity to climate change, including the racial disparities conferred upon the American Indian population in the region, the low median home values, the high percentage of homes receiving some form of public assistance, and a higher degree of natural-resource-dependent economies.
Although the Twin Ports are recognized as a regional hub for jobs and trade, poverty rates across St. Louis (MN) and Douglas (WI) counties, where the Twin Ports are located, remain as high or higher than the overall state poverty rates. Poverty rates across the two counties and in the Twin Ports cities are higher than their respective state levels, a statistical finding that is common when comparing urban areas to rural areas. Twenty-one percent of families with children under 18 in the City of Superior live in poverty, compared with 17 percent in Douglas County and 14 percent in the state of Wisconsin. In Duluth, 17 percent of families live in poverty, compared with 16 percent in St. Louis County and 11 percent in Minnesota. Across the region, a larger proportion of people of color live in poverty than does the population that identifies as white.
In the Twin Ports region, an individual with a low education level is more likely to live in poverty compared with the state populations. In the City of Superior, 30 percent of individuals who are not high school graduates live in poverty, compared with 24 percent in the state of Wisconsin, and in the City of Duluth, 35 percent live in poverty, compared to 25 percent in the state of Minnesota.
Biological Communities and Biocultural Indicators
I. Biotic Habitats
A freshwater estuary consists of the wetland and aquatic habitats of a large river where it flows into a Great Lake; this estuary extends from the Great Lake shores upstream along the river and its tributaries as far as the influence of the lake seiche. For the St. Louis River estuary, the lake seiche influence extends about 13 miles upstream to the area near the Highway 23 bridge in the Fond du Lac neighborhood.
Written by Carol Reschke
Carol Reschke is an ecologist emeritus, US Environmental Protection Agency
In the NERRS Habitat Classification System (Kutcher 2008), Great Lakes estuaries like the Lake Superior Reserve are not included in the “Estuarine” system, because the Estuarine system is defined as tidal wetlands and aquatic habitats with water levels influenced by ocean tides. This definition restricts habitats included in the Estuarine system to marine coastal estuaries. Though the wetlands and aquatic habitats where a river flows into a Great Lake are defined as an estuary with irregular water level fluctuations caused by lake seiche, these fluctuations are different from ocean tides. Great Lakes estuaries like the St. Louis River system are distinct from freshwater tidal wetlands, where the water levels in the freshwater wetlands are driven by daily ocean tides with often dramatic water level fluctuations. For example, the mean tidal range in the upper Hudson River is 1.56 m (5 feet) change in water levels daily (Yozzo et al., 2005). Seiche water level fluctuations are typically much more subtle, averaging around 0.15 m (6 inches) in the St. Louis River estuary, except during a big storm.
Using the NERRS Classification System, the biotic habitats occurring in the Reserve are classified into five systems: Riverine Habitats, Palustrine Habitats, Lacustrine Habitats, Upland Habitats, and Cultural Land Cover. Riverine habitats include all aquatic habitats contained within a river channel except wetlands dominated by trees, shrubs, or persistent emergent plants. Wetlands dominated by trees, shrubs, or emergents are classified as Palustrine. Nearly all of the Riverine aquatic habitats along the St. Louis River, Pokegama River, Red River, Nemadji River, and the Duluth-Superior harbor within the Reserve boundaries are classified as Lower Perennial subsystem, which means the gradient or slope of the river is low, water velocity is slow, and some water flows throughout the year. The substrate consists mainly of sand and mud, and the floodplain is well developed (Kutcher 2008). Based on data summaries in the Lake Superior Streams website (LakeSuperiorStreams 2009), the maximum summer water temperatures in the estuary range from ca 20°C to 27°C which falls midway between what the Wisconsin DNR defines as “cold” (less than 22°C) and “warm” (greater than 25°C) in their list of Aquatic Types. The water alkalinity of the St. Louis River in the estuary varies from ca 25 ppm to ca 90 ppm, which also straddles the definitions of “soft” (total alkalinity less than 50 ppm) and “hard” (total alkalinity greater than 50 ppm) in the WDNR list of Aquatic Types.
There are likely some small areas of Upper Perennial subsystem in smaller tributaries, especially in the Red River Breaks area and upper reaches of Pokegama River, including Pokegama Carnegie Wetlands State Natural Area (SNA). Aquatic habitats in Allouez Bay that are either unvegetated or dominated by submerged aquatic vegetation and lacking any emergent or floating-leaved vegetation, are classified as Riverine habitats.
Palustrine habitats include vegetated wetlands dominated by emergent and floating-leaved herbs, shrubs, and/or trees. Palustrine habitats are common along the main channels of the St. Louis River and the mouths of larger tributaries, as well as in wet depressions inland. In the Reserve the Palustrine Habitats are subdivided into two subsystems: Perennial Water subsystem and Intermittent or Saturated subsystem. Aquatic beds dominated by submerged aquatic plants are in the Perennial Water subsystem, and wetlands with emergents, shrubs, or trees are in the Intermittent or Saturated subsystem.
Lacustrine habitats, as defined in the NERRS Classification System, are much less common in the Reserve. Aquatic habitats in Lake Superior along the NE shore of Wisconsin Point are Lacustrine habitats, they are the unvegetated sandy shores of Lake Superior Whereas, the aquatic habitats along the shores of Allouez Bay on the SW shore of Wisconsin Point are Riverine habitats.
Upland habitats are all the dry, well-drained lands within the Reserve including upland forests, shrublands, and grasslands with natural vegetation. All of the Upland Habitats in the Reserve are within the Inland Upland subsystem, and the classes are defined by dominant vegetation type, or in sparsely vegetated areas by substrate.
The Cultural Land Cover system was developed as a way to identify areas significantly modified by humans, such as developed lands and agricultural lands; there are only a few small areas of Cultural Land Cover habitats within the Reserve. Each of the five habitat systems is divided into subsystems, classes, and subclasses.
II. Microbiological Components
Solving Mysteries with Microbes in the St. Louis River Estuary
Though we cannot easily see it, microbial life has an undeniably significant importance in the ecological communities around us. Microbes play a crucial role in material cycling within aquatic environments, mutually influencing lower trophic levels like benthic organisms and aquatic macrophytes, water quality, and public health. Understanding microbial assemblages and function begins with examination of field sediment, water, and tissue samples (see Figure 16).
By Kelly Duhn and Dr. Chan Lan Chun
Kelly Duhn is a Water Resources Science Graduate Student, at the University of Minnesota Duluth & Dr. Chan Lan Chun is an Associate Professor in the Engineering Department, University of Minnesota Duluth.
Using microbiological methods such as culturing, enumeration, and molecular techniques such as DNA sequencing and polymerase chain reaction, we can develop a baseline understanding of microbial community composition and structure. Then, when compared alongside physiochemical and other biological data, we can better understand ecological patterns and connections associated with water quality. Microbial research benefits our understanding of the conditions that may drive ecosystem health and services in the Saint Louis River Estuary.
A microbiome in the estuary is analogous to what is found in humans as structural community compositions directly affect system functionality. The microbiome varies with seasonal dynamics, climate change, and anthropogenic activities. Particularly, anthropogenic impacts, primarily in the form of terrestrial runoff, wastewater outfall, and water displacement from ship ballasts during docking, also contribute to physicochemical regimes, such as nutrient and sediment loading and other harmful chemicals. Moreover, international shipping trade may also influence invasive species dispersal, which can carry with them new pathogens. Importantly, the pollutants and their associated changes influence microbial assemblages in the estuary. Such impacts may drive a decrease in diversity, often coinciding with increased pathogens. As a major component at the base of aquatic food webs, the estuary microbiota plasticity eventually manifests in higher trophic levels, such as invertebrates and fish, affecting recreational uses and ecological resilience of the estuary (Hintze et al., 2017).
To date, little is known about the diversity and assemblages of microorganisms and their inferred functionality in the St. Louis River Estuary. With advances of next-generation sequencing technologies with traditional microbiology methods, we start to shed light on baseline variation of microbial communities in the estuary within certain habitats. A recent study has examined microbial communities associated with sediment and emergent plants in wild rice (Zizania palustris) beds. The wild rice has ecological functions and cultural value in the Great Lakes regions, but distribution and abundance has been reduced across the range, including the St. Louis River estuary (MNDNR 2008, Myrbo et al., 2017). Soil-microbe interactions influence plant communities and may be especially important in wild rice persistence and reestablishment.
Characterization of the microbial communities in natural wild rice habitats have shown that nitrogen cycling genes are more abundant in the root tissues (Figure 17) at self-sustaining sites and point to archaeal methanogens as being key players in nitrogen fixation (McMillian 2021). When evaluating the variation in phylogenetic structure with physiochemical parameters and macrophyte composition, self-sustaining wild rice beds were relatively oligotrophic with limited nitrogen, suggesting importance of nitrogen fixers. These findings suggest microbial composition is important to the fitness of wild rice populations and has the potential to beneficially influence future management actions and decisions. Restoration of wild rice wetlands will improve long- term protection of native species and aquatic biodiversity, as well as support management of this culturally and ecologically important natural resource.
Characterization of the estuary microbiota also provide benefits to assess recreational water quality and public health in the estuary. It is well known that fecal contamination of surface waters is a widespread environmental problem and can be a risk to public health as well as result in economic and recreational losses. Conventionally, E. coli and enterococcus are currently used as indicators to predict the presence and degree of fecal pollution and potential pathogens in water (Atlas et al., 1991), as the method to detect individual pathogens can be physically laborious and costly. However, the quantitative measurement of the indicators is the basis for risk assessment studies and Total Maximum Daily Load determination but does not indicate the sources of fecal pollution responsible for impairment. Over a decade, researchers have developed a sequencing-based Microbial Source Tracking (MST) methodology in the Duluth Harbor and St. Louis River estuary to accurately determine sources of fecal pollution (Brown et al., 2017 and DeGuire 2021). This methodology sets out to match DNA sequences from a catalog of known animal feces to those recovered in environmental samples to determine sources of fecal pollution. The studies using source tracking have identified contributors to pollution, which can then be recognized and addressed by developing effective control strategies. Frequent contributors to fecal contamination in the estuary include dogs, geese and humans.
As this sequencing-based source tracking approach uses whole bacterial communities rather than certain bacterial species, it has the resolving power to uncover undiscovered diversity and microbial functions, extending its application to determine potential contributors beyond fecal contamination. For example, bacterial community characterization with the source tracking method has been applied to determine the influence of ballast water discharge on aquatic communities and establish a basic understanding of potential pathogenic threats to humans and fishes within various locations in the estuary (Zimmer, 2022). This research has helped distinguish between exposure to legacy contaminants and novel contamination which may be temporary in nature. Better understanding of wildlife pathogen transfer from ballast water and its community variance and persistence through ranging physiochemical conditions may help in harbor maintenance decisions, such as dredging, as community changes in the sediment after exposure can be long lasting.

Figure 16: Initial collection of field sediment, water, and tissue samples for laboratory examination. Credit: Kelly Dunn, University of Minnesota Duluth.

Figure 17: Roots of a wild rice plant containing nitrogen cycling genes. Credit: Kelly Dunn, University of Minnesota Duluth.
In addition to these microbiota research, this environmental DNA (eDNA) method can be a valuable tool to invasive species detection. Invasive species monitoring has also been traditionally laborious and expensive, often consisting of netting, trapping, and scuba diving. Sampling estuary water and examining presence and quantity of target species through eDNA analysis may provide a modern, less time-intensive approach to wildlife and other macroinvertebrate sampling. By comparing eDNA quantification results to traditional sampling methods, sampling protocols can be adapted for improved accuracy, which may be especially important to monitoring highly mobile species populations.
Just as animal species migrate, congregate, and reproduce seasonally, so does life too small to be seen with the naked eye. Through examining the microbial assemblages of bacteria, fungi, and archaea that exist temporarily in sediments and the water column, we can gain better insight into ecosystem functions and how these microbial communities may impact the success of other plant and animal populations.
III. Plankton
The Duluth-Superior Harbor is one of the major bulk cargo ports in the Laurentian Great Lakes. The overall magnitude of shipping and deballasting activity in the Duluth-Superior Harbor makes the location a likely hub for the arrival (invasion) and export (donation) of aquatic biota within the Great Lakes, including non-native taxa. Recent species inventories of benthic and pelagic invertebrates in Lake Superior and the Duluth-Superior Harbor and St. Louis River Estuary were done by Grigorovich et al., (2003) and Trebitz et al., (2010).
Excerpted and adapted from: Defining the Risk-Release Relationship of Daphnia magna, a Non-native Aquatic Species in the Environment of Duluth-Superior Harbor (2015), by Dr. Matthew TenEyck, Director, UW-Superior Lake Superior Research Institute.
A species inventory of zooplankton in the Duluth-Superior Harbor and St. Louis River Estuary was performed by TenEyck (2015) between April 2007 - April 2008. Prior, the latest quantitative open-water surveys of crustacean zooplankton were conducted by Balcer (1978) and Hargis (1984). The survey conducted by Tenyck (2015) described spatial and temporal patterns in the density and diversity of crustacean zooplankton in the Duluth-Superior Harbor and St. Louis River Estuary, analyzed the zooplankton population patterns in the context of spatial and temporal patterns in temperature, chlorophyll and other environmental factors, and identified probable windows of high and low establishment opportunity for putative zooplankton invaders based on local community measures of zooplankton density and diversity, physiochemical conditions, and arrival times of propagules in ballast water.
In the study, the Duluth-Superior Harbor and St. Louis River Estuary (hereafter DSHSLRE) were divided into three regions defined by shipping activity (Figure 18), and are coded as upstream (region 1), inner harbor (region 2), and outer harbor (region 3). Cargo ships are active in regions 2 and 3 but not in region 1. The upper reaches of the study area (region 1) retain relatively unmodified coastal and submerged habitat that includes intact wetlands and forested riparian zones. By contrast, the lower reaches (regions 2 and 3) consist of large portions of dredged river bottom and a substantial amount of residential and commercial shoreline development. Region 1 stations were the shallowest, warmest, most productive (Chl concentration), and least stable (most lotic-like) while region 3 stations were the coolest, least productive, least turbid (lowest turbidity, highest Secchi transparency), and most stable (most lentic-like) of the three regions.
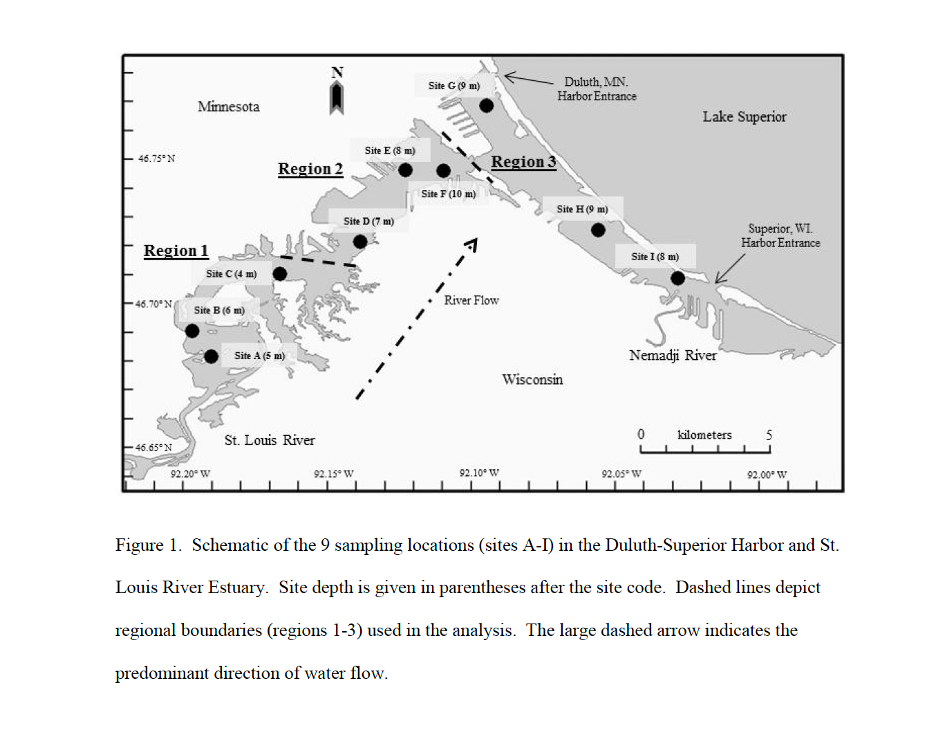
Figure 18: Figure showing 9 sampling locations in the Duluth-Superior Harbor and St. Louis River Estuary.
Among the 49 distinct taxa that were identified, approximately half at the genus level were Copepoda (Calanoida and Cyclopoida) and half were Cladocera (Table 3). Regional trends were evident in the Copepoda (Table 3). For example, Eurytemora affinis, Leptodiaptomus siciloides, and Skistodiaptomus oregonensis were present at all sites while Leptodiaptomus ashlandi and Leptodiaptomus minutus were present only at sites H and G, respectively (Table 3). Acanthocyclops vernalis, Diacyclops thomasi, Mesocyclops edax, Microcyclops rubellus, and Tropocyclops prasinus mexicanus were present at all sites, while species of the genera Eucyclops and Macrocyclops were never detected in region 3 and species of the genus Paracyclops were detected only in region 1 (Table 4).
Although about half of all Cladocera taxa were present in all three regions, several species were region specific. For example, Daphnia mendotae never appeared in region 1, and Pleuroxus, Sida crystallina, and Simocephalus never appeared in region 3 (Table 4). In contrast to region 1, the crustacean zooplankton in regions 2 and 3 more strongly reflect the species composition in Lake Superior (Balcer et al., 1984, Brown and Branstrator 2004).
Lake Superior is overwhelmingly dominated by calanoid copepods, particularly species of the genus Leptodiaptomus (Watson and Wilson 1978; Megard 1997; Zhou et al., 2001; Brown and Branstrator 2004). In Lake Superior, cladoceran zooplankton (e.g., Bosmina, Daphnia) are also common but occur predominantly seasonally and in relation to warmer water temperature (Watson and Wilson 1978). It is possible the occurrence of Lake Superior zooplankton found in regions 2 and 3 resulted from a mass of Lake Superior water mixing with DSHSLRE in regions 2 and 3 (Hoffman et al., 2010) thus creating a zooplankton community that resembles both areas (Balcer 1978).
There was also a signal in the data that suggests species richness is greater in region 1, which is upstream of Spirit Lake (sites A-C) than region 3 (contiguous with Lake Superior, sites G-I). Species richness in region 2 was typically higher than in region 3; in only two cases did region 2 richness fall below region 3. This pattern in species richness may reflect exchange with Lake Superior’s assemblage of zooplankton through the Duluth and Superior entries. Consistent with these factors, the region 1 assemblage of crustacean zooplankton was dominated by littoral genera including Camptocercus, Macrothrix, Monospilus, Pleuroxus, Sida, and Simocephalus. While the study sites for this region were located in the pelagic-like main river channel with water depths from 4-6 m, large and relatively shallow embayments (e.g., Pokegama Bay) that contained both submerged and emergent vegetation may have provided source habitat for some taxa in this littoral-rich assemblage. Two genera of Cyclopoida, Eucyclops and Macrocyclops, that were found in regions 1 and 2 but not region 3, prefer shallow littoral habitats (Balcer 1978) consistent with the littoral nature of region 1. Research by Walseng et al., (2006) suggested species richness is typically higher in littoral regions than pelagic regions due to greater habitat heterogeneity, which potentially creates higher niche opportunities. This may also be the case in DSHSLRE as suggested by the higher richness in region 1.
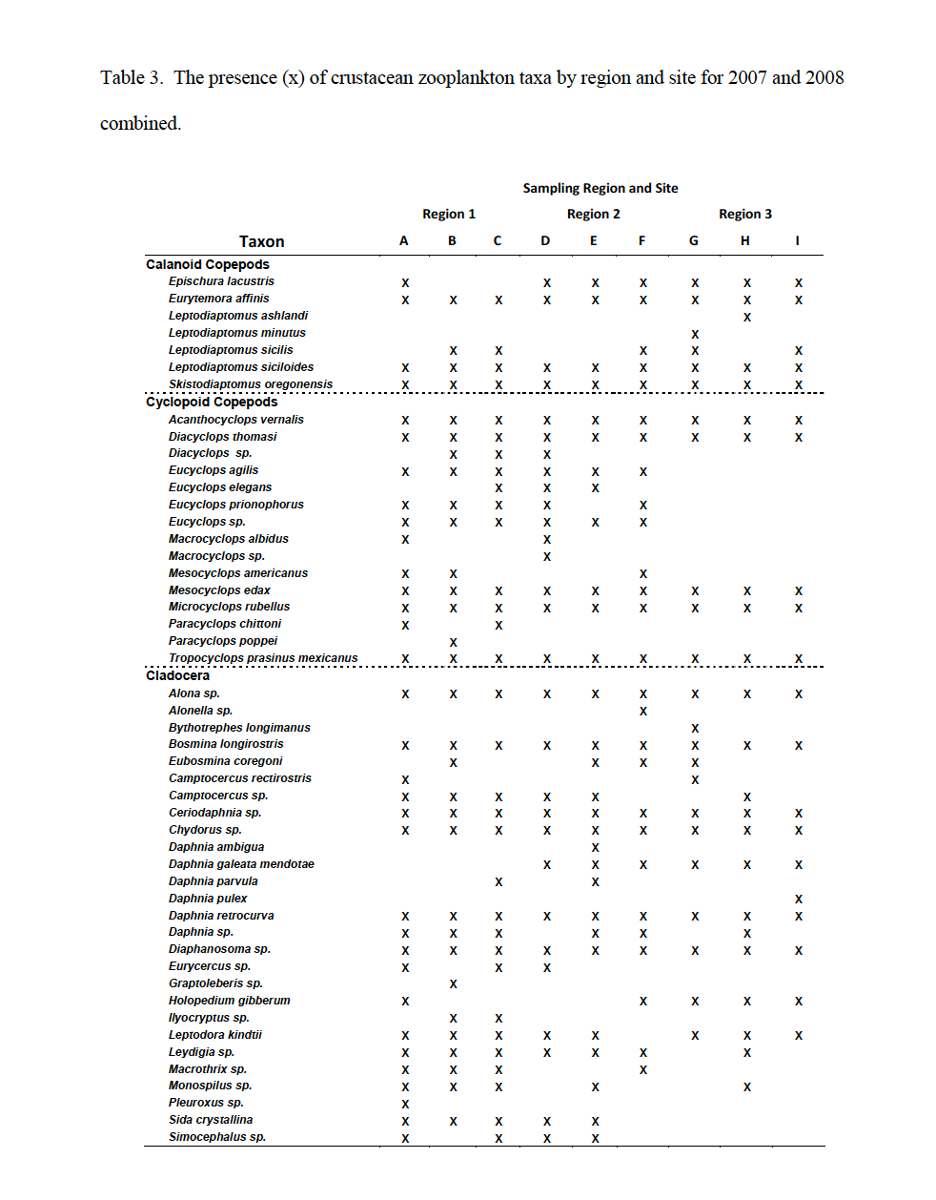
Table 3: The presence (x) of crustacean zooplankton taxa by region and site for 2007 and 2008 combined. Credit: Mathew TenEyck, UW-Superior Lake Superior Research Institute.
To summarize, there were notable spatial and temporal patterns in the density and diversity of crustacean zooplankton in the DSHSLRE. Patterns of crustacean-zooplankton growth closely reflected water temperatures and were likely responsible for the negative relationship observed with the chlorophyll concentrations. Yearly and regional variations showed that the DSHSLRE is not a static environment; physiochemical and biological conditions within region 1 changed more dramatically than regions 2 and 3 from year to year and within a given year. While each of the individual and biological processes described above played a role in identifying probable windows of high and low establishment opportunity for zooplankton invaders it is ultimately the interaction of these events that will likely lead to high establishment opportunities (Davis et al., 2005).
IV. Vegetation
Contemporary satellite images of the St. Louis River estuary are contrasted with aerial photographs from 1939 and selected areas of the 1861 Hearding map of the estuary, in many cases showing loss of emergent wetlands over time.
Vegetation and Biotic Communities of the Reserve
Historical vegetation in the Reserve area was reconstructed based on notes and maps from the Public Land Surveys, which were conducted from 1853 to 1865 (Finley 1976, Staffen et al., 2014). These land surveys were conducted to establish a grid for land ownership, not to describe vegetation; however surveyors identified witness tree species at grid corners, and sometimes would describe areas they crossed. In the mid-1800’s most of the Reserve was dominated by a distinct type of boreal forest dominated by conifers mixed with some hardwoods.
Written by Carol Reschke
Carol Reschke is an ecologist emeritus, US Environmental Protection Agency
Common trees were white pine, white spruce, paper birch, northern white cedar, aspen, tamarack, and balsam fir. Small open marshy areas were likely present inland at Pokegama Carnegie Wetlands SNA where tamarack was a common species. The extensive wetland vegetation along the St. Louis River was not recorded by land surveyors; however historical accounts by early explorers suggest the area contained extensive emergent marshes, floating bogs, and wild rice beds (SLRCAC 2002). Logging of the forests began in 1860, peaked in the 1890’s, and declined by the 1920’s (Staffen and O’Connor 2014a); the extensive logging caused significant changes to the forest and wetland vegetation. During the peak logging era, logs were floated down the St. Louis River to mills in the estuary. Upstream about a half mile from the northwest lobe of the Reserve is a large hydroelectric dam, which began operations in 1907, changing the hydrology of the wetlands in the estuary.
Vegetation in the state of Wisconsin was classified and described in detail by John Curtis, professor of plant ecology at the University of Wisconsin-Madison, and published in his book Vegetation of Wisconsin (Curtis 1959), based on data collected by Curtis, his colleagues and his students in the 1950’s. In the 1980’s the Wisconsin DNR started programs to inventory of Wisconsin’s biota and natural communities, including the Scientific Areas program and the Natural Heritage Inventory program. More recent descriptions of Wisconsin’s vegetation were published online in 2015 by the Natural Heritage Inventory program in the book Ecological Landscapes of Wisconsin (WDNR 2015), drawing heavily on the work of Curtis as well as on recent surveys conducted by DNR staff. In that 2015 publication, they define a “natural community” as an assemblage of interacting plants, animals, other organisms, and the physical environment in which they occur that is shaped primarily by natural processes and may be repeated across a landscape where similar environmental conditions prevail.” Detailed descriptions of the natural communities of Wisconsin are provided in chapter 7 of the DNR’s online publication (Epstein 2017).
The following description of vegetation in the Reserve uses the natural community nomenclature and classification presented by Epstein (2017). The communities are often named for their dominant plant species (e.g. wild rice marsh, alder thicket), a prominent environmental feature (e.g. Great Lakes Dune), or some combination of these factors. Natural communities can range in size from less than an acre to thousands of acres (Epstein 2017). Each of these natural communities are nested within a system, subsystem, class and subclass of the NERRS Habitat Classification system, as shown in Table 2.
Vegetation within half a mile of St. Louis River shoreline within the St. Louis River estuary was mapped in 2002 by Eric Epstein and Carol Reschke as part of the Lower St. Louis River Habitat Plan (SLRCAC 2002). Local studies of vegetation in portions of the Reserve were conducted by Hlina and Anderson (2009), Roesler (2013), Staffen and O’Connor (2014a, 2014b), O’Connor et al., (2018), Reschke and Hill (2020), and Hartsock et al., (2022). Vegetation monitoring data have also been collected by Reserve staff from the Sentinel Site in wetlands along the Pokegama River.
Wetland or Palustrine Natural Communities
The Reserve landscape is characterized by poorly drained, heavy red clay soils that support wetland vegetation, even on some sites with relatively higher elevations (WDNR 2015b). The most common wetland cover types in the Reserve area are marshes, sedge meadows, shrub swamps, and hardwood swamps. Some of these red clay wetlands, such as those in Pokegama Carnegie SNA are remarkable for the unusual flora they support, including several rare species: floating marsh marigold (Caltha natans), neat spike-rush (Eleocharis nitida), large-leaved avens (Geum macrophyllum), Vasey’s rush (Juncus vaseyi), sweet colt’s-foot (Petasites sagittatus), small yellow water crowfoot (Ranunculus gmelinii), tea-leaved willow (Salix planifolia), and clustered bur-reed (Sparganium glomeratum) (Hlina and Anderson 2009). The natural communities mentioned here (community names in bold italics) are described in detail by Epstein (2017) and Staffen and O’Connor (2014a, 2014b). Within the St. Louis River estuary, there is a gradient from deeper water marshes or Submergent Marsh closer to the main river channels, then shallower, somewhat protected areas with Floating-leaved Marsh, to Emergent Marshes along the river shore or on shallow sandbars. These marshes often occur in partially enclosed embayments with natural levees on the upstream end that are open to the river on the downstream ends.
The most common aquatic plants in the Submergent Marsh are water celery (Vallisneria americana), bushy pondweed (Najas flexilis), and clasping leaf pondweed (Potamogeton richardsonii). The Floating-leaved Marsh has a mix of submerged and floating-leaf plants. The most visible plants of the Floating-leaved Marsh are water lilies (Nymphaea odorata, Nuphar variegata) and floating leaf pondweed (Potamogeton natans). H Submerged plants are actually more abundant than floating-leaf plants in the Floating-leaved Marsh, and common submerged plants are common coontail (Ceratophyllum demersum), and water celery (Vallisneria americana).




Emergent Marshes in the estuary are variable, depending on the water depth and exposure to wind, waves, and water currents (Reschke and Hill 2020). The first variant, in the more exposed areas with somewhat deeper water depths (ca 0.5 to 1.4 m) the Emergent Marsh often has a hemi-marsh structure with a roughly equal ratio of open water patches interspersed with patches of emergent marsh. This hemi-marsh has submerged and floating-leaf plants common in both the open water and emergent patches. Common emergent plants in the hemi-marsh areas are sessile-fruited arrowhead (Sagittaria rigida), soft stem bulrush (Schoenoplectus tabernaemontani), and giant bur-reed (Sparganium eurycarpum). The most common submerged plants in the hemi-marsh are bushy pondweed (Najas flexilis), water celery (Vallisneria americana), and common coontail (Ceratophyllum demersum). A second variant of Emergent Marsh occurs in shoreline areas more protected from wind and waves, and at shallower water depths (ca 0.3 to 1.2 m), where the Emergent Marsh is a more continuous stand of emergents dominated by either cattails or bur-reeds and bulrushes. Most of the cattails currently found in these emergent marshes are non-native invasive cattails (Typha angustifolia and T. X glauca), and current restoration efforts in parts of the estuary are attempting to reduce or remove the often dense cover of these invasive cattails. On shallow sandbars and in some less disturbed shoreline areas, giant bur-reed (Sparganium eurycarpum), soft stem bulrush (Schoenoplectus tabernaemontani) and river bulrush (Bolboschoenus fluviatilis) are common emergents. Some of the more disturbed shoreline Emergent Marshes also have invasive purple loosestrife (Lythrum salicaria). Most of these Emergent Marshes include a mix of submerged and floating leaf plants in between and under the emergent plants, with common coontail (Ceratophyllum demersum) and greater duckweed (Spirodela polyrrhiza) the most common.
Wild Rice Marsh is less common in the estuary, found mainly along the shores of Pokegama River and in the Fond du Lac marshes, including North Duck Hunter Bay and Walleye Alley Bay following restoration efforts (Vogt 2021). Wild Rice Marsh is dominated by manoomin or wild rice (Zizania palustris), and in the estuary there are usually a few other aquatic species present, such as common bladderwort (Utricularia vulgaris), common coontail (Ceratophyllum demersum), and sessile-fruited arrowhead (Sagittaria rigida). Wild rice or manoomin is an annual plant with varying densities depending on water levels and disturbances, and they are especially vulnerable to browsing by geese. Wild Rice Marshes in the estuary have been monitored annually since 2015 by the 1854 Treaty Authority (Vogt 2021).
The large wetland complex at Allouez Bay has areas of Northern Sedge Meadow and Great Lakes Shore Fen, and there are smaller areas of Northern Sedge Meadow along Pokegama River. The Northern Sedge Meadows are dominated by sedges, with Carex lacustris and C. stricta the most common. Great Lakes Shore Fens are peatlands where the vegetation usually forms an extensive floating mat composed mostly of sedges with Carex lasiocarpa usually the dominant sedge, and sweet gale (Myrica gale) a common low shrub. Collectively all these marshes and wetlands are described as Great Lakes coastal wetlands (Hartsock et al., 2022) or Great Lakes Estuary complex (Epstein 2017).
There are patches of shrub swamps scattered around the Reserve, including Alder Thickets with dense stands of speckled alder (Alnus incana) that are common along the main channel of the St. Louis River, and Shrub-carr dominated by willows (Salix spp.) and dogwoods (Cornus spp.) scattered throughout the Reserve area. Some hardwood swamps and alder thickets have small openings of Northern Sedge Meadows in high-elevation clay flats with a high water table.
In the upstream reaches of the St. Louis River there are hardwood swamps present on sandbars and in flats along the river, most common is Northern Hardwood Swamp dominated by black ash (Fraxinus nigra), but there are also patches of Floodplain Forest with balsam poplar (Populus balsamifera), green ash (Fraxinus pensylvanica), and bur oak (Quercus macrocarpa). Black ash forests are being rapidly reduced by the invasive emerald ash borer, which was detected in Superior in 2013. Small areas within upland forests along the river shores include shaded seepage areas called Forested Seeps. In poorly-drained areas with woody peats or muck, there are some small stands of Northern Wet-Mesic Forest with northern white cedar (Thuja occidentalis), tamarack (Larix laricina), black spruce (Picea mariana), white spruce (Picea glauca) and balsam fir (Picea abies).
Upland Natural Communities
The red lacustrine clay plain so characteristic of the Reserve has been carved and eroded by the Red River, Pokegama River, and numerous smaller tributaries, creating many deeply incised ravines. These ravines can be more than 150 feet deep, with Boreal Forest on the slopes of the ravines (Staffen et al., 2014). The Boreal Forest of the ravine slopes are dominated by large white pine (Pinus strobus), mixed with white spruce (Picea glauca) and sometimes northern white cedar (Thuja occidentalis). Much of the level to rolling forested upland beyond the ravines have Aspen-Birch Forests dominated by trembling aspen (Populus tremuloides) with some paper birch (Betula papyrifera) and occasional conifers in the understory. These Aspen-Birch Forests are early successional forests that became established following logging. Poorly drained trembling aspen stands have an understory of speckled alder, and grade into shrub swamps and black ash swamps. In areas with mesic or moist soils, there are small stands of Northern Mesic Forest dominated by sugar maple (Acer sachharum), yellow birch (Betula alleghaniensis), red oak (Quercus rubra) and basswood (Tilia americana).
The northwestern peninsula of the Reserve, just east of the Fond du Lac Dam, has a sandstone bedrock substrate, and slopes rise steeply from the river. These slopes are partially forested with Dry-mesic Forest with white pine (Pinus strobus), red pine (P. resinosa), northern white cedar (Thuja occidentalis) most abundant.
Along the steep bluffs of red clay ravines in the Red River Breaks area, and on river shore bluffs along the St. Louis River there are eroding or slumping clay slopes that are characteristic of the red lacustrine clay plain; this natural community is called Clay Seepage Bluff (Epstein 2017). The plant assemblages on these eroding bluffs are highly variable, and may range from steep woods, to patches of scattered trees and shrubs, to unstable and almost bare slopes with a few forbs (Epstein 2017). Both native and exotic weedy pioneer plants such as fireweed (Epilobium angustifolium) and orange hawkweed (Hieracium aurantiacum) are found on the Clay Seepage Bluffs.
On the Wisconsin Point sandspit there are three natural communities on the sandy shoreline, with mostly unvegetated Great Lakes Beach along the Lake Superior shore, then Great Lake Dune occurs just inland from the beach, with American beach-grass (Ammophila breviligulata) and beach pea (Lathyrus japonicus) the most common plants. And further inland on the sandspit is Northern Dry Forest dominated by white pine (Pinus strobus).
Significance of Plants in the Anishinaabe Culture
Anishinaabe people are a group of Indigenous peoples within the United States and Canada, including the Ojibwe, Potawatomi, and Ottawa tribes living near Lake Superior. Knowledge of plants is an essential part of traditional Anishinaabe culture, and familiarity with local plants is essential for providing food, traditional lodging, medicines, and materials for cultural practices. The natural environment in the Ojibwe language is referred to as “Manidoo ogitigaan” or Great Spirit’s garden (Great Lakes Wild Rice Initiative 2020), and this garden includes all the human persons, plants, animals, rocks and other things found in a landscape (Davidson-Hunt et al., 2005). In the Anishinaabe language, plants, animals, rocks, and some items made from them are considered to be animate, like persons, and they are spoken of with the same grammar as human persons (David et al., 2019). It is generally understood that plants are to be respected as relatives, and that respect is inherent in the language (Kimmerer 2013). There is also a reciprocity in the traditional relationship with plants, with established protocols for gathering plants, such as giving a gift of tobacco to the plant (or to the land or water) before a plant is gathered for any purpose (Kimmerer 2013, David et al., 2019). This reciprocity was explained by elders from Shoal Lake First Nation (in northwest Ontario and eastern Manitoba) as an understanding that Great Spirit’s garden provides Anishinaabe people with all the things that they need to survive following a traditional Anishinaabe way of life (Davidson-Hunt et al., 2005). The relationship between Anishinaabe people and their land or environment is both material and spiritual. In return for the gifts provided by the land, plants, and animals to Anishinaabe people, there is a reciprocal responsibility expected of Anishinaabe people to take good care of the garden, to be good stewards or custodians of the natural abundance (Davidson-Hunt et al., 2005).
In studies of Ojibwe or Anishinaabe uses of plants, there are hundreds of plants in the Great Lakes area that are known to be used by Anishinaabe people (Meeker et al., 1993, Densmore 2005, Davidson-Hunt et al., 2005, Belcourt 2007). Many plant species found within the Reserve are gathered and used by Anishinaabe people for a variety of purposes. The plants described below are just a few of the most significant and most commonly used plants. Common uses are described here for plant species that occur within the Reserve.
Manoomin or wild rice (Zizania palustris) is especially significant in the Anishinaabe culture spiritually, as a food source, and as an economic resource. The Anishinaabe migration story is a legend that explains the movement of Anishinaabe people starting more than a thousand years ago from the Atlantic Northeast westward to the Great Lakes area. The migration story starts with one of seven prophets telling the people to head west to land where “food grows on the water”, and this sacred food growing on the water is called manoomin in Ojibwe or wild rice (David et al., 2019). The migration legend describes seven major stopping places where Anishinaabe people stopped and lived for a while before continuing on their journey and reaching their final destination on Madeline Island in the Apostle Islands, WI (Benton-Benai 1985). The sixth stopping place was in the St. Louis River estuary, at Spirit Island in Duluth. Manoomin became a staple in the Anishinaabe diet, and an essential part of the Anishinaabe culture, economy, and identity (Great Lakes Wild Rice Initiative 2020).
Maple trees (Acer spp., especially Acer saccharum) are another important food source; maples are called ininaatig in Ojibwe. Mature maple trees are tapped in early spring, sap is gathered and boiled down to produce syrup and sugar. Some other common food plant species that grow in the Reserve include hazelnuts (Corylus cornuta), wild leeks (Allium tricoccum), wild ginger (Asarum canadense), chokecherries (Prunus virginiana), and raspberries (Rubus spp.) (Densmore 2005, Wrobel 2020). Many species of plants that grow in the Reserve are used to make teas and medicines; some examples are northern white cedar (Thuja occidentalis), red osier dogwood or red willow (Cornus sericea), stinging nettle (Urtica dioica), sweetflag (Acorus americanus), and small yellow water lily (Nuphar variegata) (Densmore 2005, Belcourt 2007).
Many trees and shrubs are harvested to provide materials for making useful items like baskets, snowshoes, cradleboards or dikanaaganan, and for building lodges for cultural gatherings (Wrobel 2020). Black ash (Fraxinus nigra) logs are pounded and split into thin strips that are used to weave baskets, and thicker sections of ash wood are steamed and bent into snowshoes, lacrosse sticks, and rollbars for cradleboards. This practice is threatened by the invasive emerald ash borer, which kills 99% of ash trees in infested areas. Northern white cedar (Thuja occidentalis) boards are used for the backboard of cradleboards, and cedar is also carved into knocking sticks for harvesting wild rice. The inner bark of basswood (Tilia americana) is used for cordage and lashing baskets together. Small diameter poles of ironwood (Ostrya virginiana) and other hardwoods are used to construct frames for lodges. The outer bark of paper birch (Betula papyrifera) is gathered in sheets in early summer and used for making baskets, canoes, and coverings for lodges. Willow (Salix spp.) and red osier dogwood (Cornus sericea) branches are bent to form rims for birch bark baskets.

Figure 19: Making bawa'iganaak (a wild rice knocker) from white cedar. Credit: Deanna Erickson, Lake Superior Reserve.
Non-native Invasive Plant Species Reported from the Reserve and Nearby Areas
Non-native invasive plant species often become established in newly disturbed areas, but they may also become established and compromise high quality natural areas. Some non-native plants are not particularly invasive, they may be present without spreading. Invasive plants tend to be aggressive and once established, they can displace native plants and in some cases, they can alter ecological processes in the habitat. Wisconsin has an invasive species rule, Wis. Admin. Code NR 40, which defines Prohibited and Restricted invasive species. Prohibited invasive species are species that are likely to spread, and may cause economic or environmental harm, but only have limited populations in the area of the state where they are Prohibited; they are species for which regional eradication or containment may be feasible. Restricted invasive species are already established in the region where they are Restricted, cause economic or environmental harm, and they are species for which regional eradication or containment may not be feasible. Three species listed as Prohibited invasive species, and seventeen species listed as Restricted invasive species have been reported from within the Reserve (Staffen and O’Connor 2014a, Staffen and O’Connor 2014b, Hartsock et al., 2022).
Clough Island has the most non-native invasive species within the Reserve, including three Prohibited species: Amur honeysuckle (Lonicera maackii), Japanese barberry (Berberis thunbergii), and common reed (Phragmites australis). There are two subspecies of common reed, and one is native in the Great Lakes area (American common reed, Phragmites australis subsp. americanus), and the other is an aggressive invasive (Phragmites australis subsp. australis). Both subspecies are present in the estuary, but there have been considerable efforts to eradicate the invasive subspecies (P. australis subsp. australis) where it was found at Grassy Point, just north of Clough Island. The common reeds reported at Clough Island were not identified to subspecies. Clough Island also has twelve Restricted species present. Restricted species in upland areas of Clough Island include common buckthorn (Rhamnus cathartica), Canada thistle (Cirsium arvense), Morrow’s honeysuckle (Lonicera morrowii), Tartarian honeysuckle (Lonicera tatarica), Bell’s honeysuckle (Lonicera X bella), Amur maple (Acer ginnala), wild parsnip (Pastinaca sativa), tansy (Tanacetum vulgare), and garden valerian (Valeriana officinalis). Restricted species in wetlands around Clough Island include purple loosestrife (Lythrum salicaria), and the two narrowleaf cattails (Typha angustifolia, Typha X glauca).
Eight Restricted species are reported from the St. Louis River and along the shores, including buckthorn, wild parsnip, tansy, and garden valerian in upland areas, and narrowleaf cattails, yellow iris (Iris pseudacorus), Eurasian water-milfoil (Myriophyllum spicatum) and curly-leaf pondweed (Potamogeton crispus) in wetlands. The two aquatic invasive plants, Eurasian water-milfoil and curly-leaf pondweed are uncommon and rarely found in the estuary near the Reserve, and they are not always found each year in the estuary.

Figure 20: Buckthorn plants growing under ash dominated forests in the upper St. Louis River estuary in 2021. Credit: Deanna Erickson, Lake Superior Reserve.
Allouez Bay has one Prohibited species: common reed (Phragmites australis), but the subspecies was not reported (Hartsock et al., 2022). Allouez Bay has five Restricted species, including the two narrowleaf cattails, purple loosestrife, yellow iris, and garden valerian.
In the Fond du Lac Islands area, three restricted species have been reported: the two narrowleaf cattail species and purple loosestrife. The native subspecies common reed (Phragmites australis subsp. americanus) is present at North Bay, just north of the Fond du Lac Islands. In the Red River Breaks area, three Restricted species have been reported: common buckthorn, leafy spurge (Euphorbia esula), and common forget-me-not (Myosotis scorpioides). In the Pokegama Carnegie SNA, three Restricted species were reported: Canada thistle, purple loosestrife, and garden valerian. And along the Pokegama River including in the Sentinel Site, three Restricted species are common: the two narrowleaf cattail species, and purple loosestrife.
V. Invertebrates
The Lake Superior National Estuary Research Reserve, along with the surrounding, impactful St. Louis River Estuary (SLRE), Nemadji River Estuary, and the Duluth-Superior Harbor, provide diverse habitats for aquatic macroinvertebrates in the western tip of Lake Superior. Important features to the invertebrates include water depth, water flow, substrate type, and levels of contaminants.
Written by Dr. Kurt Schmude
Dr. Kurt Schmude is a Professor and Senior Scientist, UW-Superior Lake Superior Research Institute. 2022.
Depths range from shallow water in nearshore and wetland areas, to as much as 30+ feet in deep slips where ships dock. Current varies from still (lentic) water to strong (lotic) current, as created by the St. Louis and Nemadji rivers, as well as waves and seiches produced by Lake Superior. Diverse substrates include sand, silt, gravel, rocks, glacial-lake clay, submerged and emergent vegetation, detritus, and contaminants (e.g., coal, taconite pellets). Combinations of these substrates are common. Levels of contaminants range from uncontaminated to very contaminated (e.g., U.S. EPA Superfund sites).
Figure 21: Sorting macroinvertebrates from the St. Louis River estuary is an important component of Reserve educational programming. Credit: Deanna Erickson, Lake Superior Reserve.
The phylum Annelida (segmented worms) is represented by at least 27 species of leeches (Hirudinea) and at least 75 species of aquatic worms, mainly the class Oligochaeta. Oligochaetes are numerous and diverse within the different substrates. Species of tolerant oligochaetes are found abundantly in organically enriched areas, while less tolerant species can be plentiful in uncontaminated areas, especially where sand is dominant. Another annelid worm is the interesting polychaete, Manayunkia speciosa. Polychaetes are mostly saltwater or brackish water inhabitants, but some freshwater species occur in the world, and the SLRE is home to this species. Numbers of this small worm can be high, especially in shallow, sandy, silty areas.
Other phyla of worms include Platyhelminthes (flatworms), Nematomorpha (horsehair worms), Nemata (nematodes or round worms), and Nemertea (proboscis worms). There is an unknown number of species for each of the first three groups because it is difficult to identify them, especially the very abundant nematodes. There is likely only one species of the uncommon Nemertea.
The Mollusca include the clams and mussels (Bivalvia), and the snails (Gastropoda). The native bivalves are represented by two groups. The mussels (Unionidae) include at least four uncommon species, and the commonly found fingernail clams (Sphaeriidae) include 25 species that range from rare to common; a few sphaeriids may be introduced species. The molluscs also include three invasive species. Corbicula fluminea (Asian clam) occurs only in discrete areas. However, the invasive zebra mussel, Dreissena polymorpha, is much more widely distributed and is abundant in some areas, especially shallower, nearshore areas, often where contamination or other disturbance is present. The invasive quagga mussel, Dreissena rostriformis, is also present, but its distribution and abundance are more limited. The gastropods (snails) are comprised of at least 32 species that range from rare to common. They are more abundant and diverse in shallower, sandy, vegetated areas. Three invasive species include the New Zealand mud snail, Potomopyrgus antipodarum, Chinese mystery snail, Cipangopaludina chinensis, and big-ear radix, Radix auricularia, but their distributions are currently limited. The invasive faucet snail, Bithynia tentaculata, is more prevalent.
The crustacean arthropods are represented by three groups. At least six species of Amphipoda (scuds), three species of Isopoda (sowbugs), and four species of Decapoda (crayfish) occur in various habitats, with most of them being common and abundant in shallower, vegetated areas. Two invasive species have been found, including the scud, Echinogammarus ischnus, and the crayfish, Faxonius rusticus, but their distributions are limited.
The diverse and abundant aquatic mites (Trombidiformes) is another group of arthropods. At least 26 genera of mites have been found, but the number of species is unknown because it is difficult to identify these diverse, tiny creatures.
The most diverse and perhaps abundant group of arthropods include the Hexapoda (insects). The Ephemeroptera (mayflies) are represented by at least 25 species in six families, most of which are lentic and occur most commonly in shallower, vegetated areas. A few species occur on rocky substrates in deeper, lotic habitats. The two most common mayflies include the large Hexagenia limbata, and the diminutive Caenis. Hexagenia can be found in a variety of habitats throughout the area, and larvae range in numbers from 0 to 100 /m², or more. This species is often used as an indicator species for good habitat and water quality. Some other Areas of Concern on the Great Lakes do not have similar populations of Hexagenia as those found in the SLRE (e.g., Bay of Green Bay in Lake Michigan). Caenis occurs widely in shallower, sandy, silty habitats.
There are at least 16 species of dragonflies and damselflies (Odonata) in six families. The larvae are found in shallower, sandy, silty areas, usually amongst vegetation.
The caddisflies (Trichoptera) are represented by at least 24 species in 8 families , with most of them found in shallower, sandy, silty areas where vegetation is present. One species, Phylocentropus placidus (Dipseudopsidae), occurs within a variety of substrates in shallow to deep water throughout the SLRE.
The Coleoptera (beetles) contain at least 47 species in 12 families. More species are likely to be discovered in the diverse families Dytiscidae, Gyrinidae, Haliplidae, and Hydrophilidae when shallower, wetland areas are more thoroughly surveyed.
The same statement is true for the aquatic bugs (Hemiptera). This group contains at least 21 species in 10 families. More thorough surveys in shallow, vegetated areas will assuredly reveal more species, especially in the speciose group Corixidae (water boatmen).
The most abundant and diverse group of insects is the Diptera (true flies). There are at least 164 taxa (genera and/or species) in 13 families that have been found, with the vast majority of them belonging to the diverse midge family Chironomidae (131+). This particular family has species that occur in very contaminated and organically enriched habitats, to areas that are predominantly undisturbed. Most species exhibit varying degrees of tolerance to disturbance.
Three less diverse insect groups include at least three species of caterpillars (Lepidoptera), and at least one species each of fishflies (Megaloptera – Sialis), and spongillaflies (Neuroptera – Spongillidae).
Finally, there are populations of miscellaneous groups, including Ectoprocta (Bryozoa – moss animals), Cnidara (Hydra and Cordylophora caspia – an introduced species), Porifera (sponges), and Tardigrada (water bears).
VI. Fish, Reptiles, and Amphibians
Fish of the St. Louis Estuary
The St. Louis River estuary is an interstate water that has 8,606 acres of littoral area with a maximum depth of approximately 55 feet. The estuary is an open system directly connected to Lake Superior with a very diverse fish community including 69 species sampled during investigations from 1980-2018.
Adapted from the Minnesota DNR Lake Management Plan for the St. Louis River estuary, 2019 and the Minnesota DNR Lake Sturgeon Management plan for the St. Louis River estuary, 2019.
The freshwater estuary and its tributaries are unusual in having such a variety of habitat types supporting a large and diverse assemblage of native fish species, many of which inhabit the near-shore waters Lake Superior and utilize the estuary for spawning and nursery purposes. Forage species such as emerald shiner, spottail shiner, log perch and johnny darters inhabit the estuary, along with piscivorous species such as yellow perch, smallmouth bass, musky, walleye, and northern pike. Lake sturgeon historically used the estuary for spawning but were likely extirpated during the mid twentieth century pollution era. A two decade multi-agency stocking program has created a sub-adult population with larval fry first documented in 2011.
Prior to 1978, no fisheries management was conducted due to water pollution and very low fishing pressure. However, water quality improved dramatically after Western Lake Superior Sanitary District began treating industrial and municipal effluent from Cloquet and Duluth in 1979.
The area from the Minnesota/Wisconsin boundary cable to the Fond du Lac Dam is managed as a permanent fish sanctuary. The sanctuary is intended to protect the primary spawning area for Walleye and Lake Sturgeon, as well as aggregations of fish below the dam year round. The multi-species spawning area located between the Highway 23 bridge and the boundary cable has been closed from the end of the gamefish season through May 18th each year since 1979.
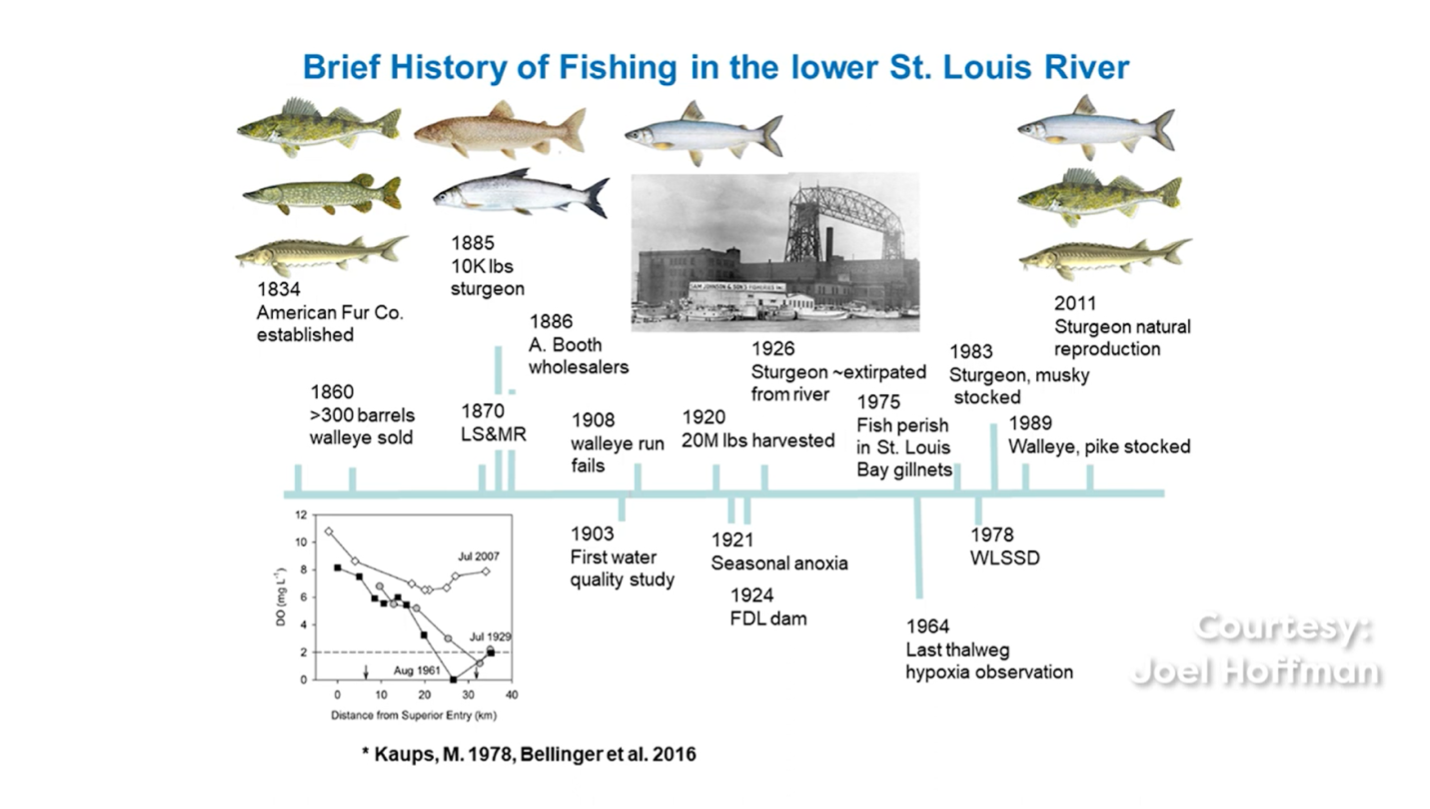
Figure 22: A brief history of fishing in the lower St. Louis River. Courtesy: Joel Hoffman. Credit: Kaups, M. 1978, Bellinger et al., 2016.
Lake Sturgeon, name in Ojibwe (pronounced nah-mey), are native to the estuary but were extirpated in the early 1900s as a result of habitat alterations, exploitation and degraded water quality. Re-introduction of lake Sturgeon in the SLRE started in the early 1980’s and has been a cooperative effort between Tribal and State agencies. The Lake Sturgeon rehabilitation plan for Lake Superior defines a self-sustaining population as one with a minimum of 1,500 mature adults, a roughly equal sex ratio, 20 or more year-classes of adult fish, evidence of natural reproduction through the collection of viable eggs, and measurable recruitment of age 0-5 fish. Fond du Lac Resource Management initiated a lake sturgeon stocking program into the Upper St. Louis River in 1998.

Figure 23: Lake Sturgeon spawn in the upper reaches of the St. Louis River Estuary. Credit: Minnesota Department of Natural Resources.
The goal of the program is to establish a self-sustaining population in the upper St. Louis River by 2042 (Howes 2015). Downstream emigration of sturgeon from the upper St. Louis River into the estuary has not been documented but is conceivable, especially for larval or juvenile sturgeon. Natural reproduction was first observed by Fond du Lac Resource Management staff below the Fond du Lac dam in 2011, but concerns about recruitment remain.
The estuary is a popular sport fishery, with walleye one of the more popular species. Walleye fry were stocked yearly in the estuary from 1979 to 2006, except 1987 and 1988, as part of a state wide requirement to stock 10% of the fry reared from the eggs taken from a river or lake. Walleye stocking was discontinued after 2006 due to the threat of Viral Hemorrhagic Septicemia (VHS). The percentage of anglers targeting walleye based on three summer creel surveys ranged from 55.6% to 68.3%. During two winter creel surveys 39.2% and 72.2% of anglers targeted walleye. Mean length of harvested walleye ranged from 17.5 to 18.5 inches during the summer and 17.4 to 18.5 inches during the winter. Tissue samples (fin clips) were collected above Highway 23 from 141 walleye during the 2015 spawning run. These tissue samples were analyzed using stable isotope analysis to determine whether a given fish was consuming forage from the estuary, Lake Superior, or both (transient). The vast majority of fish samples, 117, were classified as transient suggesting that these fish spend time foraging in both the estuary and Lake Superior. This analysis also found that Lake Superior resident fish were significantly longer than transient fish, which were significantly longer than estuary resident walleye.
Muskellunge are also a popular sport fish. Muskellunge stocking has been a cooperative effort between agencies. A total of 91,846 Muskellunge fingerlings and 4,231 yearlings were stocked from 1985 through 2005 and 2015. Muskellunge harvest was initially managed with a bag limit of one and minimum length for harvest of 36 inches. The length regulation was changed to a minimum length for harvest of 40 inches in the mid 1990's. In an effort to maintain consistent fisheries regulations between the two states, Minnesota increased the minimum length for harvest of Muskellunge to 50 inches in 2007 to match the Wisconsin regulation. Large-framed trapnet assessments of spawning adult Muskellunge from 1997 to 2018 have shown mean and maximum lengths increasing over time though young-of-year muskellunge growth rates are slower than those found in Northern Wisconsin lakes. Angler collected tissue samples, fin clips, have also been used to monitor muskellunge movement. Results from samples analyzed in 2015 found that 21 % of the Muskellunge sampled consumed at least some forage from Lake Superior (Hoffman 2016).
Other fish stocked in the estuary include smallmouth bass (1981, 1984, 1985) and black crappie (1983, 1985, 1986 and 1988-90), bluegill (1981), brown trout (1985-1992) and northern pike (1989-1993). Yellow perch have not been stocked, and have a bag limit of 25 fish with no size minimum. Channel Catfish have not been stocked and the re-population of the estuary with Channel Catfish after water quality improved in 1978 was likely a result of downstream migration from the St. Louis River above Cloquet. Bullheads, white sucker and redhorse have declined in numbers since the 1980s and 90s.
Exotic fish in the estuary include Eurasian ruffe, alewife, brook silverside, round goby, tubenose goby, threespine stickleback, white perch, white bass, sea lamprey, rainbow smelt, and common carp. Most of these exotic species have not been effectively sampled by experimental gillnets. Gizzard shad have also been identified in the estuary but have not been documented in assessment gear. Anglers have reported that catch rates for White Perch have increased, and gillnet catch rates have increased slightly. The estuary has also been identified as a potential spawning area for Sea Lamprey.
Maintenance of commercial shipping activity results in deep navigation channels being maintained by dredging. This significantly reduces productive littoral habitat within the estuary. Historically, spoils from dredging operations have also been used to fill wetlands surrounding the estuary. Dredging and commercial activities have dramatically altered bottom bathymetry, distribution of aquatic plants and overall littoral productivity within the estuary. The loss of open water is a concern in the estuary. Alterations to the runway configuration at the Sky Harbor Airport filled 7.6 acres of open water in 2019 and there have also been inquiries into filling other areas to expand piers and parking.
Climate change has the potential to affect fish populations in the estuary. A changing climate can alter fish behavior, distribution, development, reproduction, and survival. A changing climate can also increase the likelihood of the estuary being impacted by additional invasive species. Because the rate and magnitude of climate change may exceed the adaptive capacity of fish species, active (adaptive) management is required to increase resilience and reduce the impacts of climate change. The primary adaptation strategy for climate change will be to maintain a resilient and healthy watershed where possible. Protecting important features such as riparian wetlands, floodplains, terraces, sediment transfer areas, water storage areas, nutrient cycling capabilities, natural hydrologic regimes, and riparian corridors will help maintain system resiliency to climate change.
Reptiles and Amphibians of the St. Louis River Estuary
Of the approximate 60 species of amphibians and reptiles collectively found in Minnesota and Wisconsin, there is reasonable support from species range maps that 23 species occur in the St. Louis River Estuary but more species may occur. There has not been scientific documentation of amphibians and reptiles’ presence or abundance for most species in the estuary.
Written by Gerald J. Niemi
Gerald J. Niemi, Professor Emeritus, Department of Biology, University of Minnesota, Duluth. 2022.
Except for toads and frogs, the secretive behavior of salamanders and several snakes and turtles are not conducive to easy identification. Correct documentation usually requires extensive searches and effort by individuals with field experience with these organisms. Both documentation and research needs exist to track amphibian and reptile populations and verify the diversity in the St. Louis River estuary.
Amphibians and reptiles, in contrast with the warm-blooded birds and mammals, are cold-blooded animals. The cooler waters of Lake Superior and its estuarine wetlands provide a harsh environment for these poikilothermic critters. Yet, with a warming climate, these conditions are in the process of change. Amphibian and reptile species found south of the St. Louis River may have opportunities for movements north. If they are not already present, several species that are found farther south in Minnesota or Wisconsin may colonize the St. Louis River ecosystem. They include salamander species like the eastern tiger, spotted, and four-toed salamander, frogs like Cope’s gray treefrog and the pickerel frog, snakes like the common water snake, or turtles like the spiny softshell.
At the present time, herpetologists are reasonably confident that the following species can be found within the boundaries of the St. Louis River ecosystem: (1) four species of salamanders (blue-spotted, eastern red-backed, mudpuppy, and eastern newt), (2) one toad (American toad), (3) eight species of frogs [gray treefrog, American bullfrog (introduced), green, northern leopard, mink, wood, spring peeper, and boreal chorus], (4) six species of snakes (ring-necked, red-bellied, and common garter, DeKay’s brownsnake, western foxsnake, and eastern hog-nosed snake), and (5) four species of turtles (snapping, painted, Blanding’s, and wood). Depending on the species and their life stage, these organisms are found in the open waters, wetlands, riparian areas, open habitats, and forests of this ecosystem.
The herpetological species of conservation concern in the St. Louis River include the Blanding’s turtle (Threatened in MN and Special Concern in WI), wood turtle (Threatened in MN and WI) and Special Concern status for the eastern red-backed salamander (MN), and mink frog (WI). The American bullfrog is a special case in the estuary. The species has been detected at Allouez Bay and along the Nemadji River. It is considered a game species in Minnesota and Wisconsin and, therefore, laws regulate how many can be captured for food (frog legs), for use as fish bait, for the pet trade, or for educational purposes. In addition, the bullfrog can be considered an invasive species when found outside its normal range of which the St. Louis River is on the northern periphery in its United States range. The bullfrog is known to displace many native amphibians once it colonizes a new area.
The most studied of the herpetological species are the toads and frogs (collectively called anurans because they are all in the scientific order Anura) and to some extent the wood turtle which has received some study because of its threatened status. In 2011 as part of the Great Lakes Restoration Initiative, a monitoring program for anurans was established in the St. Louis River and the program is planned to continue through 2025. During their spring mating season, anurans are relatively easy to identify by their vocalizations. A total of 18 sites have been sampled within the St. Louis River with frequency of sampling ranging from once to eight annual samples. Annual samples include three visits during early spring, late spring, and early summer. The visits are timed so the potential species present at a site can be detected when they are vocalizing. Seasonal timing is based on temperature thresholds of water and air because temperature controls when each of the anurans are calling. All nine species of anurans that are known to occur in the St. Louis River have been detected using this sampling method. Seven species are relatively ubiquitous because they are found at many or most of the sites in the St. Louis River. These species include the American toad, gray treefrog, spring peeper, boreal chorus frog, green frog, northern leopard frog, and wood frog, while the bullfrog and mink frog are relatively rare and have only been found at a few sites.
The anuran monitoring project of the Great Lakes Restoration Initiative extends throughout the entire Great Lakes from the St. Louis River in Minnesota and Wisconsin to the St. Lawrence River at the eastern end of Lake Ontario in the state of New York. The Initiative included both the United States and Canada. The standardized and coordinated methodology for gathering anuran data has allowed the development of ways to both measure the ecological condition of the anuran community and now, as the program approaches over ten years, trends in their populations can be estimated.
Within the NERR boundaries, wetlands at Allouez Bay, Dwight’s Point, Clough Island, Kimball’s Bay, and the Pokegama River have been sampled in the St. Louis River. Unfortunately, because these anurans need to be counted at night, there are many safety concerns and access prohibits routine anuran sampling for many wetlands in the St. Louis River. Many of the wetlands, especially in the western end of the St. Louis River estuary (e.g., Red River breaks), have not been sampled but provide potentially suitable habitat to house many species of toads, frogs, and likely other amphibian and reptilian species.
In summary, the St. Louis River ecosystem potentially has 23 species or more of amphibians and reptiles. Besides toads and frogs, most of the documented information about the herpetological species of the St. Louis River are derived from basic inventories or incidental observations from the public. There is a need for a systematic inventory of the salamanders, turtles, and snakes of the St. Louis River as well as surveys of the anurans in the many inaccessible wetlands by trained and experienced field personnel. All of these species provide essential ecosystem services (e.g., population control such as insectivory or seed dispersal) to the long-term sustainability and ecological health of the St. Louis River.
Sounds of the frogs on Spirit Island
Play the audio file below to hear the sound of the frogs of Spirit Island
VII. Birds and Mammals
Birds of the St. Louis River Estuary
The St. Louis River Estuary is one of the best places to observe birds in the states of Minnesota and Wisconsin. The geographic location of this massive freshwater system at the far western end of Lake Superior is a mecca for attracting an amazing mixture of birds. Birds tend not to fly over large bodies of water like Lake Superior, the largest lake in the world based on surface area. During northward or southward migrations, birds move along the coastline, often in large numbers, and especially through Minnesota and Wisconsin Point.
Written by Gerald J. Niemi
Gerald J. Niemi is a Professor Emeritus at the University of Minnesota Duluth
Over 300 bird species have been observed in the myriad of wetlands, coastlines, open water, forests, and its human infrastructure. Spectacular spring and fall migrations of loons, waterfowl, grebes, shorebirds, gulls, raptors, and songbirds are routinely observed here. Examples include thousands of scaup (Greater and Lesser), Bonaparte’s Gulls, Bald Eagles (over 5,000 counted in spring of 2018 and 2021), Broad-winged Hawks, Common Nighthawks, and numerous songbirds like Blue Jays and American Robins. Many rare birds of the upper Midwest can be found here such as the Minnesota endangered Horned Grebe (100s during migration) and the Wisconsin endangered Red-necked Grebe. The list of rare birds that have been observed in the estuary is exceedingly long. Examples include the Harlequin Duck, three species of scoters (Surf, White-winged, and Black), Red-throated and Pacific Loon, Great Egret, Mississippi Kite, Gyrfalcon, Yellow Rail, American Avocet, Willet, Ruddy Turnstone, Iceland Gull, Ivory Gull, Scissor-tailed Flycatcher, and an endless number of songbirds such as the Blue-gray Gnatcatcher, American Pipit, or Rusty Blackbird. In some winters, more than a dozen Snowy Owls can be observed. Sightings for many of these species attract hundreds of bird watchers and nature enthusiasts. Wisconsin Point even has a Jaegerfest in September when up to three species of jaegers (Pomarine, Long-tailed, or Parasitic) can be observed, all rare species in the United States from the far north.
This estuary provides important breeding habitat for several species of conservation concern in Minnesota or Wisconsin. The Common Tern (Threatened in MN; Endangered in WI) nests on Interstate Island in the Duluth-Superior Harbor and is only one of two colonies on Lake Superior. The Piping Plover (Endangered in the US, MN, and WI) formerly nested in the St. Louis River but has been extirpated since the 1980s; despite continued efforts in habitat management to re-attract them, only a few migratory individuals have been detected.

Figure 24: Common tern nesting sites on Interstate Island are monitored throughout the summer season. Fencing is designed to reduce predation by gulls. Credit: Minnesota Department of Natural Resources.
This estuary provides important breeding habitat for several species of conservation concern in Minnesota or Wisconsin. The Common Tern (Threatened in MN; Endangered in WI) nests on Interstate Island in the Duluth-Superior Harbor and is only one of two colonies on Lake Superior. The Piping Plover (Endangered in the US, MN, and WI) formerly nested in the St. Louis River but has been extirpated since the 1980s; despite continued efforts in habitat management to re-attract them, only a few migratory individuals have been detected. They nest in isolated sandy-cobble beach habitat that also are attractive to humans, their pets, and many predators. Several pairs of Peregrine Falcon (Special Concern–MN, Endangered-WI) nest on human-made structures like bridges, buildings, or large smokestacks and the once federally-endangered Bald Eagle now thrives with several nesting on large trees throughout this ecosystem. The exquisite Black Tern (Endangered-WI) formerly nested at Allouez Bay and near Morgan Park in MN in the 1970s. Efforts are underway to again attract this species to the extensive wetlands of the St. Louis River.
Other species that occur during the breeding season with Special Concern conservation status include the Trumpeter Swan (MN), American Bittern (WI), Least Bittern (WI), American Woodcock (WI), Common Nighthawk (WI), Least Flycatcher (WI), Ruby-crowned Kinglet (WI), Swainson’s Thrush (WI), and Brewer’s Blackbird (WI). The wetlands and uplands throughout the St. Louis River provide habitat for many of these species as well as large populations of Sedge Wrens, Marsh Wrens, and Alder Flycatchers.
Breeding and migratory birds can be observed anywhere in this extensive river system from the downtown areas of Duluth and Superior to the popular and accessible Minnesota and Wisconsin Points, or to the more remote forests of the Superior Municipal Forest or Chambers Grove in the Fond du Lac community. Minnesota and Wisconsin Points can be exciting places to observe birds during migration. On foggy, cool mornings in spring these areas can be dripping with birds, especially colorful spring warblers and other songbirds. These conditions create fallouts that ground northbound bird migrants as they encounter severe weather when approaching Lake Superior.
Other popular areas to view birds include the Duluth Lake walk, the Waaabizheshikana Trail (formerly the Western Waterfront Trail) at the western end of Duluth, and the Osaugie Trail in Superior; each follow closely along the Lake Superior or the St. Louis River shoreline. The vast wetlands of the St. Louis River are places not to be missed such as those at Allouez Bay, along the Nemadji or Pokegama Rivers, and those surrounding the Oliver Bridge in Wisconsin. In Minnesota, the larger wetlands are best accessed at Indian Point, Mud Lake, surrounding Erie Pier, and at the recently restored Radio Tower Bay in the Gary-New Duluth neighborhood.
Within the physical boundaries of the Lake Superior National Estuarine Research Reserve, the Wisconsin Point-Allouez Bay area is an ornithological gem. Almost any bird species found in the entire St. Louis River system, has likely occurred in this area. It has boundless potential as breeding habitat for many bird species of the region and its wetlands for the Trumpeter Swan, Black Tern, American and Least Bittern, Horned Grebe, Sora, Virginia Rail, and many others. The habitats along the Nemadji River, Pokegama River, and especially the intricate matrix of islands and channels from the Oliver Bridge west to the Hwy 23 Bridge offer much to be explored. These areas provide superb conditions for nesting Bald Eagles, Least Bittern, Spotted Sandpipers, and Belted Kingfishers as well as for many other riparian and wetland bird species like bitterns, herons, waterfowl, swallows, and numerous songbirds. Many of the forested areas from the Superior Municipal Forest to the Mont du Lac region have a distinct boreal influence because of the abundance of coniferous trees like white cedar, tamarack, black and white spruce, and pines. Common boreal-type bird species found nesting in these forests include Common Raven, Hermit and Swainson’s Thrush, Magnolia Warbler, Blackburnian Warbler, and Canada Warbler.
The mid-continent geography and habitats of the St. Louis River estuary are a magnet for attracting a wide variety of migratory, wintering, and breeding birds. It also is unique because it is situated among several human population centers and, therefore, substantial challenges have and will continue to occur within the area to balance future development, conservation, and preservation. Thoughtful planning and effective leadership will be essential to maintain the integrity and long-term sustainability of this exceptional bird habitat.
MAMMALS
The St. Louis River estuary is home to many species of mammals, although limited research and monitoring has been done on this taxonomic group. The water and wetlands of the estuary provide habitat for several semi-aquatic species, such as river otters, muskrats, beavers, and mink. An assessment of these semi-aquatic mammals in the St. Louis River Estuary was done by the Wisconsin Department of Natural Resources in partnership with the University of Wisconsin – Madison from 2014 – 2016.
By Morgan Swingen
Morgan Swingen is a wildlife biologist with the 1854 Treaty Authority

Figure 25: A River Otter (Lutra canadensis) captured on a wildlife camera near the estuary (phto credit: Bryn Evans)
This research documented the presence of all four species throughout the estuary. Beavers and muskrats both feed on leaves, stems, and roots of aquatic vegetation, and beavers also consume woody plants such as aspen, willow, birch, and maple. River otters and mink are carnivores, feeding mainly on fish, crustaceans, amphibians, reptiles, small mammals, bird eggs, and small birds. As high trophic-level predators, both mink and river otters have been identified as potential indicator species in which PCBs and heavy metals present in impaired waters may bioaccumulate.
The lands of the Reserve include a large amount of upland forest which has been largely un-surveyed for mammals but likely provides habitat for a number of generalist species such as white-tailed deer, coyotes, red and gray fox, bobcats, raccoons, black bears, striped skunks, porcupines, weasels, snowshoe hare, eastern cottontails, woodchucks, and squirrels.
The Red River Breaks parcel has been identified as having southernmost remnant sections of boreal forest in the U.S. However, mammalian species normally associated with boreal forests such as moose and lynx are unlikely to occur on Reserve lands due to the fragmented nature of this habitat type in this area.
Wolves were extirpated from Wisconsin around 1960 but have re-populated the northern part of the state after protections were put in place under the Endangered Species Act in the 1970s. Natural re-population occurred as wolves dispersed from populations in neighboring northeastern Minnesota. Reserve lands could provide good habitat for wolves, especially in areas with less human activity and high white-tailed deer populations, such as the Red River Breaks parcel of the St. Louis River Streambank Protection Area.

Figure 26: Beaver (Castor canadensis) change shoreline habitats in the estuary (phto credit: Deanna Erickson)
Recent monitoring using radio-collars has shown wolf use of the Reserve lands in recent years, although it is unknown if there are any resident wolf packs on the Reserve. Wolves are currently listed as federally endangered in Wisconsin under the Endangered Species Act, although the status has changed multiple times in the last decade.
Terrestrial furbearers such as fisher and American marten may be present on Reserve lands, but have not been confirmed by any scientific surveys. The American marten is more closely associated with closed-canopy coniferous forests, which are limited within the Reserve, and are therefore likely uncommon. Fishers require large trees for denning, but can use deciduous trees when conifers are scarce. Both American marten and fisher have been recorded in Jay Cooke State Park just west of the Reserve in Minnesota. American martens are listed as state-endangered in Wisconsin, and fishers were once rare but populations have rebounded since re-introduction efforts in the 1980s and 1990s.
Small mammals (mice, shrews, voles, and moles) are an important component of the Reserve ecosystem, especially as prey for birds and larger mammals. Previous small mammal assessment work on and adjacent to current Reserve lands have confirmed the presence of common species such as the white-footed mouse, deer mouse, short-tailed shrew, masked shrew, meadow vole, and meadow jumping mouse. Other small mammals likely present within the Reserve include the red-backed vole, woodland jumping mouse, star-nosed mole, and American water shrew. This area of Wisconsin is also home to at least six species of squirrels, including tree squirrels and ground squirrels. Common tree squirrels include the gray squirrel, red squirrel, and northern flying squirrel, and ground squirrels include the chipmunk, thirteen-lined ground squirrel, and the Franklin’s ground squirrel, which is a species of Special Concern in Wisconsin.
The geographic ranges of seven species of bats also overlap with the NERR lands, although little research has been done in this area. Acoustic surveys conducted along the St. Louis River in 2013 confirmed calls from little brown bats and big brown bats, which were the most common species of bats at that time. White-Nose Syndrome (WNS), a fungal disease which causes high levels of mortality in some bat species, was first detected in Wisconsin in 2014 and in Minnesota in 2016. Little brown bats, northern long-eared bats, and tricolored bats have been greatly impacted by this disease and their numbers have declined greatly in recent years in both Wisconsin and Minnesota. Current population levels of the various bat species in the Reserve area are therefore unknown, but it is likely there are currently higher proportions of big brown bats and migratory bats such as eastern red bats.
| Scientific Name | Common Name |
|---|---|
| Odocoileus virginianus | White-tailed deer |
| Canis lupus | Gray wolf |
| Canis latrans | Coyote |
| Urocyon cenereoargenteus | Gray fox |
| Vulpes vulpes | Red fox |
| Lynx rufus | Bobcat |
| Ursus americanus | American black bear |
| Mephitis mephitis | Striped skunk |
| Procyon lotor | Raccoon |
| Martes americana | American marten |
| Pekania pennanti | Fisher |
| Lontra canadensis | River otter |
| Neovision vison | American mink |
| Mustela frenata | Long-tailed weasel |
| Mustela erminea | Short-tailed weasel |
| Castor canadensis | Beaver |
| Erethizon dorsatum | North American porcupine |
| Sciurus carolinensis | Eastern gray squirrel |
| Tamiasciurus hudsonicus | Red squirrel |
| Glaucomys sabrinus | Northern flying squirrel |
| Tamias striatus | Eastern chipmunk |
| Ictidomys tridecemlineatus | Thirteen-lined ground squirrel |
| Poliocitellus franklinii | Franklin’s ground squirrel |
| Marmota monax | Woodchuck |
| Sylvilagus floridanus | Eastern cottontail |
| Lepus americanus | Snowshoe hare |
| Myotis lucifugus | Little brown bat |
| Myotis septentrionalis | Northern long-eared bat |
| Eptesicus fuscus | Big brown bat |
| Lasionycteris noctivagans | Silver-haired bat |
| Lasiurus borealis | Eastern red bat |
| Lasiurus cinereus | Hoary bat |
| Perimyotis Subflavus | Tricolored Bat |
| Ondatra zibethicus | Muskrat |
| Peromyscus leucopus | White-footed mouse |
| Peromyscus maniculatus | Deer mouse |
| Microtus pennsylvanicus | Meadow vole |
| Myodes gapperi | Southern red-backed vole |
| Napaeozapus insignis | Woodland jumping mouse |
| Zapus hudsonius | Meadow jumping mouse |
| Sorex cinereus | Masked shrew |
| Sorex palustris | American water shrew |
| Blarina brevicauda | Short-tailed shrew |
| Condylura cristata | Star-nosed mole |
Ecological Setting and Conditions
I. Origin and Evolution of the Estuary
The geologic history of the Lower St. Louis River can be reconstructed from the rocks and sediments exposed in the river bed and along the shoreline. The present St. Louis River channel was shaped primarily by the glaciers of the Pleistocene epoch, which began advancing and retreating approximately 2 million years ago and receded for the last time around 10,000 years ago. Left behind was a complex pattern of sediment, including moraines, drumlins, beach sands, and lake-bottom clays.
Excerpted and adapted from: the Final Environmental Impact Statement for the Lake Superior National Estuarine Research Reserve and various reports produced by the Lake Superior Reserve, as cited.
The landscape of the Lower St. Louis River area today is a nearly level lake plain of lacustrine clays. The bedrock is mostly sandstones and shale covered by water-reworked moraine, forming mostly well-drained soils. The bedrock over which the St. Louis River flows is part of the Canadian Shield, the ancient core of the North American continent. The red-clay, sand, and silt cover the bedrock on the Wisconsin side of the river; remnants of a time when the region was covered by the Glacial Lake Duluth, which was formed by the melt-water trapped in front of the retreating Superior Lobe of the Laurentide ice sheet. Some basalts occur in the northern part of this region as a result of continental rifting that occurred over one million years ago.
At the end of the most recent Ice Age, massive amounts of ice as thick as several hundred feet retreated from much of the Great Lakes Basin. Post glacial rebound, also known as isostatic rebound, is occurring more rapidly along the northeastern and eastern portions of Lake Superior causing uplift in the earth’s crust that “tilts” the Basin toward the southwest, thereby flooding lake water into river mouths along the southwestern shore, creating the drowned river mouth system of the St. Louis River estuary.
Totalling nine miles in length, the baymouth bars of Wisconsin and Minnesota Point that establish the estuarine character of the river mouth were formed by deposition of quartz sand eroded by wind and wave action from the Lake Superior south shore (Loy 1963). Sands mixed with some North Shore gravel till built across the mouth of the river from the Superior South Shore to the North Shore in present day Duluth over the past 3000-4000 years. The force of water from the Nemadji River subsequently created an opening in the sandbar, establishing the two Points. An important constructive force of the bay mouth bar complex is wave action, especially generated by northeast winds, but wind erosion and deposition on the back dune and wave action in Superior Bay are also influential and may play a larger role as wind speed and frequencies increase in the context of climate change.

Figure 27: A painting by Ojibwe Finnish artist Carl Gawboy on display at the Superior Public Museum shows the creation of the river outflow between Wisconsin and Minnesota Points as the work of an otter. (Courtesy Superior Public Library)
The United States Geological Survey has found that water levels in the southwestern portions of Lake Superior have risen approximately 15 to 18 feet over the past 2000 years. USGS estimates that the lake level rise in those areas is occurring at a rate of one inch per decade. The hydrology of the St. Louis River is determined by a combination of both surface water and groundwater. Groundwater feeds the St. Louis River headwaters, the Seven Beavers/Sand Lake peatlands systems. Despite some early attempts to drain the peatlands and wetlands, much of the peatlands are still present. The wetland areas receive significant contributions from groundwater and contribute to the base flow of the St. Louis River. Surface water runoff from the St. Louis River watershed is the primary source of freshwater into the estuarine system.
II. Influence of Physical Environment on the Biota
The St. Louis River watershed is dominated by woody wetlands (~46%) followed by forested area (~29%) with only approximately 3.5% being developed (includes high intensity, low intensity, and open space, C-CAP Land Cover Atlas). The surrounding landscape of the estuary transitions from highly forested in the upper reaches to the more industrialized Duluth-Superior Twin Ports region at the mouth, where it flows into Lake Superior. The estuary itself is 24 km long with a surface area of ~50 km2. The upper boundary of the estuary is approximately defined by the most upstream location influenced by the seiche in Lake Superior (near the Fond du Lac Dam), resulting in bi-directional hydrologic exchange between the SLRE and the lake multiple times per day (Trebitz 2006). The river mouth is located between the baymouth bars of Wisconsin and Minnesota Point, constricting flow and allowing significant mixing of lake and river water. The hydrology of the estuary is complex, with river channels of varying size, back bays, islands, and wetlands. The estuary, including the Nemadji River, primarily flows through a highly erodible, phosphorus-rich red clay plain (Stortz et al., 1976). Modeling studies have shown that shoreline erosion and riverine inputs from the Nemadji river are the biggest contributors of sediment and nutrients in the western arm of the lake (Robertson 1997). The estimated mean daily discharge in the thalweg near the mouth of the estuary is approximately 73 m/s (Bellinger et al., 2016), but flow throughout the estuary is highly variable.
Despite significant reductions in point-source pollution, phosphorus (P) concentrations have remained elevated, and more recently, toxic cyanobacterial blooms have been observed in the estuary and in coastal Lake Superior along the south shore from the Duluth-Superior Twin ports region to the Apostle Islands (Sterner et al., 2020). Previous work on cyanobacterial blooms and nutrient loading in coastal Lake Superior has focused on relatively small tributaries along the south shore (Reinl et al., 2020b), but the estuary, a major input to Lake Superior and heavily impacted by human activities, has not been given equal consideration as a contributor to the HABs in the lake. Paleolimnological studies have also revealed an increase in cyanobacterial pigment in the SLRE since the 1990s (Alexson et al. 2018)
Ice conditions are also influential on biota and productivity in the estuary. A 2020 Reserve-led survey of late-winter under-ice water quality in Perch Lake, Mud Lake and Pokegama Bays revealed three consistent patterns with respect to dissolved oxygen concentrations: 1) DO was, on average, relatively low, 2) DO is lower at the bottom than at the middle of the water column, and 3) DO decreases with increasing distance from hydrologic connections to the main channel of the estuary (Bockenstadt et al., 2020). These findings are consistent with previous research (Garono et al., 2019), and demonstrate that DO concentrations in estuarine back bays remain low immediately prior to ice-out. Ice out on the St. Louis River in the 2020 study occurred during the week of March 30th. The decreasing dissolved oxygen associated with increasing distance from hydrologic connections to the estuary indicate the potential importance of water exchange to increase dissolved oxygen and viable winter habitats for fish within the bays.
The St. Louis River is impacted by methylmercury contamination and mercury in the water column interacts with sediments and associated bacteria to transform into methylmercury, which bioaccumulates in the food web. Fish consumption advisories related to mercury are in place in the estuary for children, women and men (MPCA 2022). In 2022, the Minnesota Pollution Control Agency was working with Wisconsin and the Fond du Lac Band of Lake Superior Ojibwe towards the development of a total maximum daily load (TMDL) derived from the identification of total mercury deposited in the watershed and the sources of methylmercury. Several studies led by Dr. Emily Graham, conducted in partnership with the Lake Superior Reserve, address trends in organic matter cycling, sediment microbiomes and methylmercury production in the St. Louis River (Graham et al., 2016; Graham et al., 2017; Graham et al., 2018).

Figure 28: Researcher Victoria Bockenstadt drills through the ice to measure water quality, including dissolved oxygen, at Perch Lake in the upper St. Louis River estuary in 2020. (Photo: Deanna Erickson, Lake Superior Reserve)

Figure 29: Credit: Deanna Erickson, Lake Superior Reserve.
Hydropower dams have altered water flow and connectivity in the St. Louis River since the first dam was constructed at Thomson in 1906. The dams, managed by Minnesota Power and all within Minnesota, are located at Knife Falls in Cloquet (completed in 1921), Scanlon (completed in 1922), Thomson and the Fond du Lac dam (completed in 1928) in far western Duluth. Limited study has been conducted on the influence and impact of the hydroelectric dams on water quality and sediment transport.
III. Biological Productivity
Coastal ecosystems and estuaries are among the most biodiverse ecosystems in the world and provide a wealth of ecosystem services (Barbier et al., 2011). These ecosystems are some of the most populated in the world because they lie at the productive interface of land and water. Since colonization and subsequent industrialization in the United States, estuaries have become severely degraded from a wide variety of issues including toxic pollutants, invasive species, eutrophication, and hypoxia. Coastal ecosystems, including estuaries, are far more studied in marine environments than in freshwater (Andersen et al., 2006b; Scavia and Bricker 2006b; Anderson et al., 2008). In fact, many definitions of estuaries state that estuaries are where freshwater rivers meet and mix with saltwater from the ocean (e.g., EPA, National Geographic Society, NOAA). However, many freshwater estuaries have analogous ecological functions to their brackish counterparts. Herdendorf (1990) outlines three major components of estuaries based on definitions for marine systems: 1) constriction at the mouth, 2) bi-directional water flow and exchange, and 3) mixing of two distinct bodies of water. The SLRE is a drowned river mouth with nine mile long baymouth bars at the outflow (Hobbs and Breckenridge 2011), resulting in constricted flow. While tides are typically less than three inches in the estuary (Herdendorf 1990), the seiche in Lake Superior results in substantial exchange between the river and the lake multiple times per day (Trebitz 2006; Smith et al., 2020). The St. Louis River estuary is biochemically distinct from Lake Superior. Lake Superior is a cold, oligotrophic lake with a mean chlorophyll-a (chl-a) concentration of 1.92 µg L-1, SRP of 0.05 µM, and NO3- of 25.81 µM (Reinl et al., 2020b). In contrast, the mean chl-a concentration in the SLRE is 9.25 ug/L, SRP is 0.58 µM, and NO3- is 14.3 µM (LSNERR SWMP data).
For more on biological productivity and community structure and processes within the Reserve, see Chapter 3c: Biological communities and biocultural indicators (LINK) or the Wisconsin Natural Heritage Inventory Program’s 2014 Rapid Ecological Assessment for the St. Louis River Streambank Protection Area and the Pokegama Carnegie Wetlands State Natural Area (WI DNR 2014).
Publications and Online Bibliography
All works cited or consulted in the creation of this site profile are listed in our online bibliography, hosted by Zotero. Zotero is a free, open-source reference management software that allows users to collect, organize, and store reference materials in online libraries. Our online bibliography can be accessed through this link: https://www.zotero.org/groups/3740017/lsnerr_site_profile_bibliography/library
No account or subscription is required to see the items stored in a zotero library.
